Christian
Churches of God
The Genetic Origin
of the Nations
(Edition 4.5
20060105-20060421-20070804-20071122-20200123)
The Bible gives a clear and definitive origin of the nations that allegedly does not accord with what modern Science is asserting. When we examine the scientific record against the historical evidence and the known movement of tribes, we find that the Bible and Science can be in accord. The problem lies in the grossly elongated mathematical models that the evolutionists have constructed based on false assumptions.
Christian
Churches of God
E-mail: secretary@ccg.org
(Copyright ã 2006, 2007, 2020 Wade Cox)
This paper may be freely copied and distributed provided it
is copied in total with no alterations or deletions. The publisher’s name and
address and the copyright notice must be included. No charge may be levied on recipients of
distributed copies. Brief quotations may
be embodied in critical articles and reviews without breaching copyright.
This paper is available from the World Wide Web page:
http://www.logon.org and http://www.ccg.org
The Genetic Origin of the Nations
One of the greatest advances in human genetics is that of measuring the DNA of both males and females. Males are measured through the Y chromosome that is handed down from father to son in a continual line that mutates over time. This is called YDNA. Only males carry this through their Y chromosome, being XY. Females have XX chromosomes. Science says this rate of mutation is much slower than the Bible demands it to be in accord with its history of the nations as descended from Noah.
The female line is measured by testing their Mitochondrial DNA (termed mtDNA) that both males and females possess. Mitochondria are grouped in Haplogroups that relate to the lines of females in which they occur and this line is passed from mother to her offspring in the X chromosome, which both male and female possess.
The YDNA and mtDNA are measured in two different ways. YDNA is measured in what are termed polymorphs. These polymorphs are allocated a numeric value and, according to the value when tested, the sub-groupings that are formed are called clades and subclades of the overall grouping which is called a Haplogroup. These values record the change in YDNA mutations and lines.
The mtDNA is determined by reporting the polymorphic site such as for example 311C, meaning a mutation has occurred at base pair 16,311 and the base that changed here was actually changed to cytosine. The number 16,000 is the commencement point for DNA numbering and thus the 16,000 is dropped and the numbers used are the numbers in excess. So 16,311 becomes 311 and the letter indicates the chemical at that point in the polymorph. It is this change of the polymorphic site that determines the genetic ancestry, as the parent passes on to the offspring the DNA polymorphisms that they have with the same or similar numerical values. When tested, these values that are not exactly the same as the parent are termed mutations. The values thus vary and have determined the tribal groupings of the world’s nations.
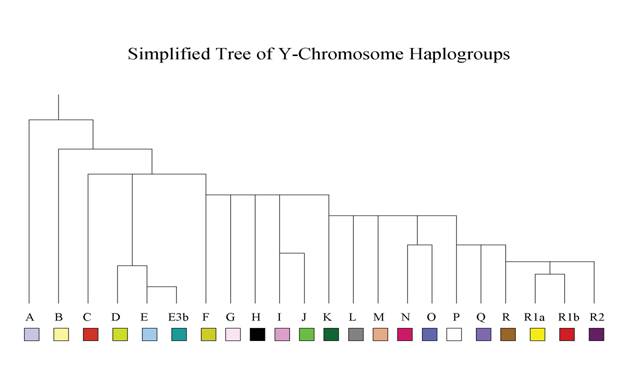
The YDNA system that has been allocated to the male human species is grouped into a series of Haplogroups from A to R. The usual extensive measurement (using the Arizona system) is usually of 37 sites as markers. Basic testing is done for the first twelve, then to 25, and then on to 37 of these polymorphic sites, or locations to determine relatedness and Haplogroup association. There could be some 100 or more markers tested for changes (a.k.a. polymorphisms). An example of what might be a typical YDNA structure is as follows:
|
1 |
393 |
x |
|
2 |
390 |
x |
|
3 |
19* |
x |
|
4 |
391 |
x |
|
5 |
385a |
x |
|
6 |
385b |
x |
|
7 |
426 |
x |
|
8 |
388 |
x |
|
9 |
439 |
x |
|
10 |
389-1 |
x |
|
11 |
392 |
x |
|
12 |
389-2 |
x |
|
13 |
458 |
x |
|
14 |
459a |
x |
|
15 |
459b |
x |
|
16 |
455 |
x |
|
17 |
454 |
x |
|
18 |
447 |
x |
|
19 |
437 |
x |
|
20 |
448 |
x |
|
21 |
449 |
x |
|
22 |
464a** |
x |
|
23 |
464b** |
x |
|
24 |
464c** |
x |
|
25 |
464d** |
x |
|
26 |
460 |
x |
|
27 |
GATA H4 |
x |
|
28 |
YCA II a |
x |
|
29 |
YCA II b |
x |
|
30 |
456 |
x |
|
31 |
607 |
x |
|
32 |
576 |
x |
|
33 |
570 |
x |
|
34 |
CDY a |
x |
|
35 |
CDY b |
x |
|
36 |
442 |
x |
|
37 |
438 |
x |
*Also known as DYS 394
**On 5/19/2003, these values were adjusted
down by 1 point because of a change in Lab nomenclature.
There are values that when tested are allocated to the right column of the table. Those values help determine whether a person has fallen into a typical Celt Haplogroup or one that is found among a Semite, or an Amerindian, or Polynesian, or Chinese or Japanese racial type. To properly determine the type, what is known as an SNP test is run, but the values from the 12, 25 and 37 marker tests point us in the right direction. No national group is of a pure genetic Haplogroup with the exception of some Amerindian tribes of the Q group in South America (from the charts produced by Dr. J. D. McDonald). The table above, depending on tested values, could be that of the typical R1b Celt of Europe or the Native Americans, both of whose DNA belongs to the last two Haplogroups on the scale.
The mitochondria, first sequenced in 1981, became known as the Cambridge Reference Sequence (CRS). The CRS has been used as a basis for comparison with individual mtDNA. In other words, any place in an individual mtDNA that has a difference from the CRS is characterized as a mutation. If a result shows no mutations at all it means that the mtDNA matches the CRS. A mutation happens when: a) a base replaces another base - for example a C (Cytosine) replaces an A (Adenine); b) a base is no longer in that position, or a deletion; and c) a new base is inserted between the other bases without replacing any other (an insertion).
That is how the mtDNA mutates. The original female DNA of the human species was confined to seven females, but in Europe only. Throughout the world there are about 26 sub-branches of the tree of humankind on the female lineages. Thus the female lineages might be argued to be explicable within the biblical terms in Europe only. The mtDNA thus requires harmonising with the Bible account by explaining the entire 26 female lines.
We are dependent on Science in its organising of the Haplogroups. As the Polymorphs mutate they alter the values and so the rate of mutation may well cause some movement between Haplogroups not yet identified.
Harmonising the
Bible and Genetics
If the Bible is to answer the challenge posed by this very important genetic discovery, then it must place this structure over the Bible record and explain the distribution of nations within terms that can take account of, and properly answer, the challenges posed by the genetic trees of both the YDNA and the mtDNA of the species.
The female lineages may well have an effect on the male, causing mutations that are not as yet known, and an examination of that system will perhaps also reveal some important aspects. However, at present, other than known recombination, no direct effect on the male mutations caused by the female is scientifically demonstrated.
However, we now know that the mitochondria are directly responsible for the mutations of the human genetic structure. It inserts into active genes and causes mutation. This is especially so in DNA that is damaged by pollution or radiation, making them susceptible to further damage from stray mitochondrial DNA. Working genes are more prone to damage as their DNA is regularly unzipped to churn out proteins (New Scientist, 18 September 2004, art. "The enemy within that targets genes", p. 11).
"Miria Richetti at the Pasteur Institute in Paris and her team identified 211 insertions of mitochondria DNA across the genome, 23 of them new, 80 percent were in genes but genes only make up 3 per cent of the genome. The targets are widely transcribed genes, probably because they have more double strand breaks, says Richetti” (ibid.).
Jef Boeke at John Hopkins University thinks that Mitochondrial DNA is unlikely to be a major source of human gene mutation. However, there seemed to be a case of this directly related to the Chernobyl radiation disaster.
By comparing the sequences of 21 ethnically diverse volunteers, Richetti's team found six insertions that were not shared by all of them. (PLoS Biology, vol. 2, e273). The scientists held that "these must have occurred after our ancestors split from the ancestors of chimps and thus could give scientists clues about the origins of ethnic groups and historical migrations" (ibid.).
We are agreed that these mitochondria are capable of mutating the human genetic structure, and the incidence of intermarriage with new mitochondria are held to cause the mutations at an accelerating rate.
That is why
isolated societies show the basic structure and less change than do the
more open societies intermingling with other mitochondrial Haplogroups. For
example, the RxR1 groups among the Australian Aborigines are also less
affected. It is not an indicator of time, but rather one of intermarriage.
The majority of the Aborignal Haplogroup C, however, is in fact C4 and is
derived from the C basic groups that spread out along the South Asian coast
from India to Vietnam. There is some C basic still there in pockets, but the
Aborigines developed into the C4 group and then subdivided into two C4
sub-groups within Australia. The Maori developed into the C2 group and the
Mongols into C3, which also went into the Americas. Both the Aboriginal and
Maori languages contain many loan words from the Malay people and indicate that
they once shared a common area and linguistic group. The Malays are, however,
predominantly Hg O but do have some other groups. The Maori left their island
home ca. 1000 CE after the rise of waters and went to New Zealand within the
Medieval Warm Period when seas were higher.
There appears to be a problem in what is termed the Y-Chromosome Phylogenetic Tree in that the Haplogroup A is the first markedly divergent fork in the tree, and thus first to diverge from Hgs B to R. All Haplogroups have a fork at which they diverge.
We will look now at an overview of the Bible account. The Bible says simply that Noah was perfect in his generations and that he begat Japheth, then Ham and then Shem. The progeny of the sons of Noah are listed and the tribes and distribution are noted and so allow us a few precious clues to measure against the DNA record.
The Bible accounts have been construed as only allowing eight people on the Ark. However, the record may refer to eight males, as females were never mentioned in ancient genealogies except in very rare instances and sometimes only in the deletion of a prefix in the father’s name (see the paper The Genealogy of the Messiah (No. 119)).
This eight-male scenario is probable as Canaan was old enough in the first wine harvest after the Flood when Noah was drunk with new wine to have either sodomised him or castrated him. The Jewish traditions discuss both scenarios. That is the reason for the biblical curse on Canaan. Some hold that it was Ham, and Canaan bore the curse rather than Ham, but that seems to be to confine the eight mouths to both the males and females. The latter case is important to the DNA record in that only the sons’ wives would have then bred on and Noah’s wife’s mtDNA would have stopped unless there were females from Noah’s wife there as well, which is possible if only males are mentioned.
The male lines were thus Japheth and Shem with their wives, and Ham and the (four) sons of Ham and their wives, totalling seven males and seven females in addition to Noah and his wife.
This would then also allow eight females including Noah’s wife, thus providing the main seven female mtDNA lines; and that seems to accord with the current findings of mtDNA Haplogroups, but for Europe only. It might be argued that some mtDNA lines are later divisions. The scientific evidence indicates that there were seven so-called “Eves” to the genetic mtDNA pool in the Caucasian but that there are 26 female lines overall. Thus, the explanation must account for the entire sequence. The argument might thus be advanced that there were 26 females on the Ark that bred on.
This view would place Science in direct harmony with the Bible record as far as female lineages are concerned. We will also find that when we look at the mtDNA record for the female lines we are then able to place the female divisions in line with the YDNA male divisions. Thus we are able to see the break-up of the families of Noah in their world distribution. We will examine this aspect later.
The Noah gene
pool
Noah was understood to be pure in his generations. The Bible also maintains that the people in the Ark were all the family of Noah. Thus, to properly account for the genetic diversity, Noah must have maintained the capacity to throw genetically distinct offspring, and this offspring had the characteristics of the line from which it came, but not the entire sequence that Noah had originally.
Conventional views of the YDNA systems are that they number back – 35,000 years and some say 60,000 to 130,000 years – and that the levels of mutation are slow. This extended view is based on mathematical models but seemingly to advance an evolutionary model. Further, no cognizance is taken of the impact of the diverse female DNA systems. The mtDNA is measured, but the effects of the female X chromosome on the male YDNA structure is considered important and it will be argued that it causes mutations in the YDNA structure at a much faster rate than the current model is believed to allow.
One of the known histories that we are able to examine is that of the Jews and we can identify a number of nations that come from them; and within their DNA structure we can see a great many people that are not actually Semite according to current wisdom. For example, 52% of all Ashkenazi Levites are not Semites according to the YDNA models and the current explanations. Thus we may have evidence here of a large number of converts to Judaism marrying into the tribes. In all probability this points to the Khazar conversion as the source of the Ashkenazi Levites. We will examine this aspect below.
Further, we know that the Buba clan of the Lemba people in Zimbabwe in the YDNA lines are all Cohenim of Levi, and the Lemba are predominantly Jewish YDNA even though they are black, having taken African wives after they were isolated at or after the Babylonian captivity. Their Judaism is much more basic than post-captivity Judaism and their calendar determination is based on the conjunction determined in advance. They have held their beliefs in a very basic form for up to 2,500 years separate from Judah.
All DNA distribution areas in this work are according to the known native populations prior to the European dispersion ca. 1500, according to testing.
For Noah to be the father of the human structure he is held to have had the capacity for the following YDNA substructure, as all humans are descended from him. Any male on the planet will have only the mutations that signify his branch and path.
M91, SRY 10831a, M42, M94, M139, M60, M181, RPS4Y711, M216, YAP M145, M203, M174, SRY 4064, M96, P29, P14, M89, M213 and the core for the subsequent derivatives.
Noah held the base YDNA that was able to mutate into these other subgroups. The divisions that sprung from him are as follows:
Sons of Ham:
YDNA Haplogroup (Hg) A: M91 and subsequent derivatives
A1: M31
A2: M6, M14, M33, M49, M71, M135, M141, P3, P4, P5, MEH1, M196, M206
A3: M32
A3a: M28, M59
A3b M144, M190,
M220
A3b1: M51
A3b2: M13, M63,
M127
A3b2a: M171
A3b2b: M118
This tree is distinct from the other Hgs B to R.
This group is found predominantly in the Sudan >45%, Ethiopia > 15%, and South Africa > 30%. Very small elements of the Hg A are found in Cameroon, seemingly in Morocco, and among the Pygmies in Central Africa. The majority of these others are ExE3b and B for the Pygmies and in South Africa, or E3b for Morocco.
Africa is predominantly ExE3b and E3b with the B group distributed among them. The incidence of A in Africa is also seen with Haplogroup C at its root, prior to a group that stayed in Africa and a branch that left Africa.
Haplogroup B is comprised of M60, M181 then:
B1: M146
B2: M182
B2a: M150
B2a1: M109, M152,
P32
B2a2: M108a
B2a2a: M43
B2b1: P6
B2b2: M115, M169
B2b3: M30, M129
B2b3a: M108b
B2b4: P7
B2b4a: P8
B2b4b: MSY2a, M211
Haplogroup B appears in smaller percentages in Mali, Cameroon, among the Pygmy of Central Africa, in South Africa, in Sudan and in Ethiopia. Haplogroups A and B do not appear in any significance outside of Africa or African Americans.
Haplogroup C is a very interesting Haplogroup. It is derived from the DNA groups M168 and P9 and then to the RPS4Y711 and M216 segments which form the basic C group. The basic C group is found among the Australian Aborigines. The divisions are:
C1: M8, M105, M131
C2: M38
C2a: P33
C3: M217, P44
C3a: M93
C3b: P39
C3c: M48, M77, M86
C4a
C4b
The C groups are found among the following people:
Australian Aborigines >65% (C4)
Maori >80% (C2)
French Polynesians >55%
Western Samoans >30%
Smaller percentages are found in Iryan Jaya, Borneo (Kalimantan), New Britain, and the Philippines;
Smaller percentages are also found in China and in Japan.
The major percentages are then found in Northern Asia:
Mongols up to 60%
Buryats up to 65%
Evenks up to 70%
Koryaks up to 35%
Altaians up to 23%
Khazaks up to 42%
Uzbeks up to 15%
Kyrgyzs some 10%
Yakuts 10%
There was an element (C3) that moved into the Amerindians in Alaska (say some 42%) and then to the Chippewa or Na-Dene in Canada (+- 6%) and the Cheyenne (>14%) and Apache (>12%) in USA. There is a very small percentage in the Greenland Eskimos showing extensive Arctic contact.
It is a matter of great significance that we have been able to unearth the point of departure for these people and tribes in Asia. The Choukoutien Upper Cave in China (discovered in 1930 and excavated by Pei 1938-1940) was found to be home to a family group occupying the same strata and comprised of Melanesoid, Eskimoid and Mongol types. Weidenreich held that the Wadjak remains were more like the adult female remains in the Upper Choukoutien Cave than with any Mediterraneans so far (R.M. and C.H. Berndt, Aboriginal Man in Australia, Angus and Robertson, 1965, pp. 30, 32-33).
Previously, Huxley (1870) had classified the Australian Aborigines as being the advance guard of the Dravidians that left the Mediterranean and Egypt and moved into India and on to Australia. He regarded the Dekkan Hill tribes, and more tenuously the Ancient Egyptians, as the only links with Aboriginal Australians (ibid., p. 33). He had classified the early men into four types: Negroids, Mongoloids, Xanthochroics and Australoids. The Tasmanian Aborigines he classified as a Melanesian branch of the Negroids. Wallace (1893) had said that the Australians were neither Negroid nor Mongoloid and classified them as early Caucasians (ibid.). However, this has been shown to be incorrect, and the Australian Aborigines are direct descendants of the division of Haplogroup C that left Africa or the Middle East. They are also comprised of another two Haplogroups, namely Hg K of the Melanesians and RxR1 basic, which is indeed a basic Aryan Haplogroup from which the R1a Slavs and the R1b Europeans are descended.
We now know from DNA that the finds in Choukoutien Cave of the same family with these three types were in fact correct. The YDNA of the tribal groups shows that these people are indeed all descended from the same paternal lines and the Australian is the basic C structure that moved into Australia from Africa via the North, with the Melanesians and Polynesians developing the Pacific, and the Mongols and what became the far-northern tribes going into Northern Asia.
There is no
published DNA of which the writer is aware of the early Australoids that links
the existing Australian Aborigines with the earlier finds in Australia. There
are two major YDNA groups in Australia and they are C4 and K Haplogroups with
an early RxR1 Haplogroup >10% which is only found in any significance in
Cameroon (>40%) in Africa, and with smaller groups in Uzbekistan and among
the Dravidians, and is rare in any event. That RxR1 group would justify the
links to Africa and the Dravidians of India. However, that is the only link
that the Australian Aborigines have with the Dravidians. The C4 group is a much
better link and so the Aborigines in the main came from the group that formed
the African groups and that also formed groups in Cameroon and North-East
Africa.
The vast
majority of Europeans are R1a and R1b and might be considered to have descended
from this earlier RxR1 DNA link. However, the RxR1 Haplogroup is considered to
preclude any ancient origin for the Aborigines. The 25% incidence of the K Hg
shows movement of the early (proto-) Melanesians into Australia from the north.
The C groups appear to have come from Africa/the Middle East into Central Asia.
This aspect is explainable biblically as the Cushite split. The Western
Cushites went into Africa through the Sudan and the Eastern Cushites went first
into India and then moved on in groups. After the Aryan invasion of India in
1000 BCE they appear to have moved out of India. The Aborigines split from the
C basic group on the South Asian coast, probably from their nomadic existence,
and moved south into Australia probably by canoe. The others moved east and
north into Asia, and then the Pacific. Thus the African origin may be explained
by a split in the Middle East rather than the move from Asia, and there is thus
a Central Asian origin not only for that element of the Aborigines but also for
the entire Mongol and Pacific systems of the C group. The R basic groups are
argued to have formed in India/Pakistan rather than being indicative of a
Middle Eastern origin with migration into Africa and Australia. We will look at
this aspect in dealing with Hg R. The diverse linguistic groups (8) for the
Aborigines indicate some eight waves into Australia for at least three tribal
Hg systems and probably more diverse migration.
There are no C
Haplogroups in South America and thus the supposed link between the Maori and
the South Americans is a myth. The predominant YDNA in America is the Q
Haplogroup, with the C coming in there in far fewer numbers and only from
Alaska into central North America in the Na-Dene, Cheyenne and Apache and on
into the Maya in Central America.
It is considered that the C group might be misallocated in the charts and perhaps should be linked closer to the K groups that form the proto-Melanesoids of Hgs K and M. It might perhaps change places with Haplogroup E, but given the A and C arguments above in Africa that might not be so. The movement of the C groups were the minor groups with Q that moved into the Americas. Conventional wisdom says they moved 12-15,000 years ago but the Bible time-frame says otherwise.
Maoris have told the writer that the traditions of the canoes, which narrate the movement of the tribes and link their genealogy to those canoes arriving in New Zealand, do in fact allow for an African origin of the Maori. However, the C grouping DNA, with the incidence of O, seems to demand the central Asian origin and movement from the Chinese coast perhaps coinciding with the expansion of the Han. However, their Malay linguistic links indicate a southward movement into the Indo-Malay archipelago where the Maori and the Aborigines split into C4 and C2 by isolation. There is a trace of East and South-East Asian (O) YDNA in the Maori but there are greater elements in the Western Samoans and, to a slightly lesser extent, among French Polynesians. This may well indicate a Malay/Philippine influence on the Pacific tribes of the C system, but such influence is not evident in Irian Jaya, Papua New Guinea, and New Britain where we might expect to find it more readily. The arrival of the Maori in New Zealand is quite late and they replaced two previous civilisations in NZ in the early centuries of the second millennium of the Current Era. The first was a Papuan-type culture, which was forced to migrate, and the latter seems to be almost Celt in its stone structures. Their movement is thus quite distinct from the earlier Australian Aboriginal movement of both C and RxR1.
This Haplogroup is formed from the group D and E at the YAP division with M145 and M203 as the stem for both and M174 breaking into the D:
D1: M15
D2: M55, M57, M64a, P37a, P41a, 12f2b
D2a: P12, P42
D2b: M116a
D2B1: M125
D2b2: M151
This Haplogroup is confined to two peoples in any significance. These are Japan (>40%) and Tibet (>50%). The incidence of the Hg indicates that the Japanese and the Tibetans once were a people occupying the Eastern Steppes and they moved north-east into Japan and south into the mountains of Tibet. The language of the Japanese is Uralic-Altaic and has much in common with Finnish and Hungarian, Turkic, Altaic, Mongolian, Manchu/Tungus, Old Korean and the Northern Siberian languages (and anciently with Basque). Thus we can assume that Haplogroup D was once a tribe with a common language system with those of Haplogroup C and others identified below. The other Hg (O) for the Japanese and Tibetan indicates that the Chinese had interbred with them over the centuries, no doubt affecting the language system and customs from the areas of their movement. The Japanese have higher incidence of Hg C than do the Tibetans, perhaps from their exposure to the Mongols, the Buryats and Koryaks. However, the incidence of the Aboriginal Ainu may also be a factor in this due to population replacement in the later Japanese invasions.
The other significant incidence of D is perhaps from the trading influence of the Japanese in early time going to Sumatra and to Malaya. The more significant incidence is in Sumatra at up to 10%. The incidence in Malaya is less than 5% as is the R1a grouping there also, and for F, C and M with a slightly higher incidence of K. Borneo has a less than 5% D group also, as do the Uygurs, Altai, the Mongols, the Kyrgyz, Uzbeks and the Siberian Eskimo.
Haplogroup E is found in Africa in the main. It continues the Yap division at M145 and M203 to SRY 4064 M96 and P29, which form E basic.
E1: M33, M132
E1a: M44
E2: M75
E2a: M41
E2b: M54, M85,
M90
E3: P2, DYS 391p
E3a: M2, P1
E3a1: M58
E3a2: M116b
E3a3: M149
E3a4: M154
E3a5: M165
E3a6: M10, M66,
M156
E3b: M35, M215 (omitted from some charts but contained in Family Tree DNA 2005 Y-Chromosome Phylogenetic Tree, see www.familytreeDNA.com)
E3b1: M78
E3b1a: M148
E3b2: M81
E3b2a: M107
E3b2b: M165
E3b3: M123
E3b3a: M34
E3b3a1: M136
The E groups are found in Africa, with smaller incidence in the Middle East and Southern Europe. Some speculate the incidence of E3a in UK to be from Roman slaves in Britain; others view them as slaves coming in later times. Both views may well be correct.
ExE3b is highest in Burkina Faso at well over 90% and perhaps 99%.
ExE3b is high in Mali (60%), Cameroon (45%), amongst the Pygmy (65%) and in South Africa (55% approx.). This group has low incidence in the Sudan and Ethiopia.
E3b is very high in Morocco (approx. 75%) with an incidence of Hg J at approx. 10%. There is significant incidence in Mali (25%), Sudan (25%), and Ethiopia (55%,) South Africa (less than 10%). 15% of Middle-East Arabs and Persians are E3b, and some 10% of Italians and Iberians, and between 5% and 10% of Germans and Russians are E3b. Up to 5% of Georgians and Armenians are also E3b. East European Jews are 25% E3b. They may well have been converted to Judaism from the Sudan and Ethiopia, as there are Semitic elements in both areas. They may have been part of the mixed multitude in the Exodus. More likely, many were the Canaanites that were spared and joined Israel after the occupation under Joshua. We will examine these aspects when we look at the Semitic Haplogroups below.
These Haplogroups A to E are the sons of Ham. We will try to identify their tribes at a later date.
As stated, Haplogroup C may well have been misallocated and probably should appear in order after where E stands in the tree, next to F and this side of what is termed the YAP divide. Both D and E are defined by an insertion on the Y that D and E share while C and F do not have this insertion. They appear together more frequently in the tribal locations.
The F Haplogroup stems from the division between the Yap M145, M203 divide for D and E and the RPS4Y 711 M216 divide for the C group. It is a small group and sometimes acts as a catchall because researchers did not do enough testing to determine the group correctly. There are small F groups in Georgia/Armenia, Persia, Uzbekistan, among the Kazan Tartar and in Khazakstan. The conclusion is that the basic root of F is nearly gone but the prolific sons survived and flourished, producing the major national groups and the mutations that flowed from that stem.
This stem determines all the other Haplogroups from F to R.
The basic F is P14, M89, M213.
In biblical terms, both Shem and Japheth passed this core Haplogroup on to all their offspring. Perhaps Ham also passed it to one his sons. The charts would make Ham’s sons widely divergent.
The mtDNA puzzle
Before we examine the YDNA Haplogroups (Hgs) further, we will pose a problem and suggest a solution.
There are 26 mtDNA Haplogroups indicating 26 female mtDNA lines. Some seven original Hgs or female “Eves” are posed for Europe. However, when we examine the tree of mtDNA we find some interesting group derivatives. The so-called “supergroups” are really only in three basic groups. In other words, they came from three main female lines. That is what we would expect to find if we assume there were only three females that bred on from the Ark, namely the wives of Shem, Ham and Japheth. These Haplogroups are all descended from a single female supergroup, namely Haplogroup L. So in reality, all females are descended from one female line, Hg L. That is super L. This line then split into L1, and then L2 and L3. The line L3 diverged and from L3 came the other mtDNA mutations. Thus, all females came from one Eve whose mtDNA line was L.
The L groups L1, L2 and L3 are all found in Africa and are the major groups almost exclusively in sub-Saharan Africa. Only from Ethiopia north do we get large diversity of the mtDNA record. That is the basic reason why evolutionists claim that we all came out of Africa. The placement of the DNA groups can be seen in the work of J. D. McDonald who has grouped them by charts of Y and mtDNA Haplogroups available at http://www.scs.uiuc.edu/~mcdonald/WorldHaplogroupsMaps.pdf.
The supergroups M and N were next to diverge or mutate. From a biblical point of view we can argue easily that L was formed with Eve and the other groups were pre-Flood divisions that came on to the Ark. Thus, we could correctly argue that L, M, and N came on to the Ark within the accepted biblical account. It is also possible that the subdivision supergroup R may have come on, depending on the number of females on the Ark. All mtDNA Haplogroups are subdivisions of L, then M and N and subsequently R, which itself is a mutation of Hg N. Thus the only argument between the Bible account and modern scientific DNA is the supposition that the mathematical models require a much longer period than the Bible chronology to mutate. That assumption is based on the premise that mtDNA does not force mutation of the Human Genome and that assumption is now being shown to be false. The Bible statement that “death came by sin” is seemingly demonstrable as a condition of the human DNA system, as we saw previously. As seen from the Pasteur Institute research, mtDNA causes mutation where damage has occurred through disease, and also radiation, as we are now discovering. We are progressively mutating in our DNA through the direct influence of the Mitochondrial DNA on the Human Genome. We started perfect with Adam and we are getting weaker over time. We did not evolve; we degenerated. Our DNA mutated with exposure to mtDNA in its variant forms. It follows as a matter of logic that diverse mtDNA must cause further mutation in the Human Genome thus affecting the Y Chromosome.
Thus, from the original Hg L we get Hg M and Hg N. Both of these groups are independent direct mutations of Hg L.
Thus, we can assume that Eve produced the line L and the three wives of Shem, Ham and Japheth are at least the three groups L, M and N. There may have been further divisions given the fact that Noah may have had daughters not mentioned and their mtDNA line may have been L, or M or N. It may have even been R, if we assume that the entire L line came in through the wife of Ham, as the L line is almost confined to the sub-Saharan tribes. We also have to address the fact that Eve was dark skinned and the fact that Adam means the one who was red. Thus the capacity for the development of skin colour was an original trait of the human creation. The explanation for the decrease in melanin, which causes pigmentation, is that the further away from the equator you go the lighter your skin must be in order to survive. You get less vitamin D with an increase in melanin. Thus, you need less capacity to absorb vitamin D where the sun is greatest, and more melanin protection, but more capacity to absorb vitamin D and less protection where the sun is decreased. So the offspring of Noah simply got lighter in melanin the further north they moved.
The wives of Shem and Japheth were of the subdivisions of L, perhaps M and N or perhaps also R.
The two groups M and N formed the following subgroups:
M produced three subdivisions:
M subgroup including
C and Z, which split from each other, and D and G;
E subdivision; and
Q subdivision
We might thus also deduce that the wives of the sons of Noah were taken from the one family lineage, maintaining purity in the generations in the female line also. The L2 and L3 split may have come from the family structure before the Flood. The daughters of Noah and the wives of the sons could have carried all three of the L subdivisions and the basic core sub-groups of M, N and perhaps R.
It is therefore possible that the women of the Ark – even if there were only the daughter of Noah and the wives of Shem, Ham and Japheth – could easily have contained the basis for the modern mtDNA diversity. We would not be surprised to find such diversity in a family of married sons even today. In Palestine, Egypt and South Pakistan it is common today to find these groups.
Supergroup M
M is found in greatest quantities and percentages among the Indians and Pakistanis. From there the group spreads north and east to the Mongols and Han Chinese and on to Japan and Taiwan. The divisions of M into C and D occurred in the Eastern steppes among the Han and the Mongols and the Japanese. The C divisions occurred among the Mongols and sub-groups. The Evenks are predominantly C with only small percentages of M, D and U. The Buryats of Central Eastern Siberia are C and D with some L3 and then the mutation G. This mutation is carried east into the Nivikhs and is most prolific amongst the Itelmen of Eastern Siberia and goes on into the Chukchi of Far Eastern Siberia.
The mutation of M into Hg E occurred among the Aboriginal Taiwanese and in Kalimantan, being prolific in Sabah. The Q variant occurred in Papua New Guinea. Thus the above mtDNA variation occurred as the tribes moved progressively east from the Hindu Cush into Mongolia, China, South-East Asia and Melanesia.
This was the original progression of the mtDNA of the M supergroup after the Flood.
Supergroup N
The supergroup N is now found mostly in Central and East Asia and among the Australian Aborigines. The N group split into:
N subgroup, which included Haplogroups I and W;
The A and X subdivision;
Y subdivision; and
R subdivision.
Concentrations of base N are among the Australian Aborigines in approximately the same distribution as the YDNA Hg C is found there. That fact indicates that these people came to Australia probably from the Eastern distribution of the Cushites and stayed isolated. Their base DNA was not mutated by other mitochondrial intrusion. Their base structure does not argue for a long occupation but rather an isolated DNA system that suffered no forced mutation by intermarriage with other mtDNA Haplogroups.
The P structure in Australia may have come in with the YDNA Hg K and RxR1 from the Melanesians. There are high concentrations of both mtDNA Hg P and Q with some M. There is a very small mtDNA M in Australia, with a similarly small percentage of mtDNA N in Papua New Guinea. This would indicate occasional exchange, probably from captive women. There is an argument that the YDNA K and the mtDNA P may have been there with the Papuans when the Tasmanian Aborigines came and in fact preceded the Australian Aboriginal YDNA C and mtDNA N. That is uncertain but the Tasmanians were of a Papuan type as were the Moriori who preceded the Maori in NZ.
The N mtDNA structure is also among the Japanese, Han Chinese, Han Taiwanese, Altai, Uzbeks, Komi, Persians, Turks and also in Southern Russia. The N supergroup split into subgroups N, I and W.
The I subgroup is somewhat rare and appears as a significant percentage, albeit a small one, among the Kurds and into Turkey, probably from the Kurds there. It is a small percentage in Egypt and then goes into Iberia, France, UK, Scandinavia and Iceland. It appears the N and I split occurred in the Middle East before the N groups went east and the I Hg went west. W is also amongst the Kurds, as is U and K and HV and the H subdivision. Thus these two divisions and the sub-groups are represented there but the parent Hg R is not represented. The N, I, and W groups are all present in Palestine/Egypt. Mitochondrial DNA Hg I probably came from the Middle East with the Anglo-Saxons or the earlier Celtic movements, perhaps even after the fall of Troy in the 11th century BCE.
The A and X mtDNA Haplogroups are predominant among the American Indians. Hg X, being a subdivision, occurred when they moved into the north-east of North America.
Y is a mutation that occurred in North Asia and is found among the Nivikhs.
Supergroup R
Haplogroup R is found among the Thai in greatest percentages, in India, South Pakistan, and in smaller percentages amongst the Han Chinese, and the Hazara.
The R supergroup split into the following:
B;
F;
HV, which split into H and V;
P;
The J and T subdivision; and
U, from which came K.
Whilst Hg R is found among the Thai and others as above, the subdivision B is more telling. B is found amongst the Han Chinese, and in greater percentages in southern China, with larger percentages among the Aboriginal Taiwanese; but it is also present among the Han Taiwanese and the Japanese. Haplogroup B moved south to Papua New Guinea and is in greatest concentrations among the Polynesians, being over 90% of their mtDNA. The B subgroup is then found across the Pacific in greatest concentrations on the west coast of North, Central and South America, indicating that the movement was from central Asia to South-East Asia to Polynesia to the Americas; but the YDNA there in the Americas is not C but Q. It is only C with Q in Alaska and then out into central North America. This C route must also have been across to the north or, YDNA Q tribes took the women of YDNA C men and wiped them out. This may be so as the recent finds of a YDNA C basic male – identified as an Australian Aborigine on the West Coast of North America and predating any other finds to date – seems to indicate this migration.
The presence of groups A, X and D together indicate a Pacific crossing from North Asia with the A division from Hg N occurring in Taiwan and Japan and spreading north to the Chukchi of Far Eastern Siberia and across the Aleutians in two groups. The northern group was to become the Alaskan Eskimos and the southern group was to become the Alaskan Amerindians. The northern groups appear to have been the primary northern migration forming the Na-Dene (Chippewa) mtDNA (predominantly from Hg A). The A group went east and north and on into Greenland, with the D groups going on into South America with A, C and B as well as into the area of what is the south-eastern USA. The Q males thus had four lines of females with them including the later mtDNA variation U being found in North America but also in the area of what is now Chile. Thus, the settlement of the Americas was late, with the YDNA and mtDNA mutations being of the second to last forms.
The HV group mutated from R in the Middle East and both R and HV are found in Persia and South Pakistan. The Hg H variant is found from the Uzbeks and the Mansi, the Hazara, and the Komi in the Central Steppes right across Europe to Iceland. H forms the major mtDNA structure of the West Europeans. It is also found in Palestine, Egypt and NW Africa to Morocco. The Hg V variant is in smaller quantities except among the Saami of northern Scandinavia where the percentages are dramatically reversed. Percentages of R subdivision J, T, U and K are found in similar distribution over Europe with the exception of the Saami and Kurds, where U is predominant, except the K variant as found among the Kurds. The K variant is absent in the Saami, indicating they did not see that variation but split from the other U groups before it occurred. The other Northern Tribes in Russia, of the Kets and the Komi, have significant U but they also have F or T variations, which the Saami do not. The Saami also have some percentage of D as well as H and some Z.
The progressive mtDNA Haplogroup divisions are shown in the attached table.

There is thus nothing in the mtDNA variations and Haplogroups to preclude the Bible story and the Genesis account being accommodated by, or accommodating, the scientific advances we find here.
Mitochondrial Haplogroups L, M, N and perhaps R were present in the women on the Ark. The mutations occurred as each group moved out from the Middle East and crossbred with moving tribes and families over time.
Next we will return to the YDNA structure to try to identify the tribes and nations involved and place them within the Genesis account.
The YDNA groups
All other YDNA groups from G to R2 are derived from one central Haplogroup F. This F group is central to both Shem’s and Japheth’s known lineages. We will start with Lineage F, which is P14, M89, M213. This line is the basic line for G, H, I, J, and K.
We know that the lineages in known Semitic nations are G, I and J. There are also some lineages of E3b African or Hamitic lineages together with some R1a and R1b. These lineages with G also spread to Turkey, Georgia/Armenia and Italy.
Conventional wisdom identifies the Middle-East Arabs as Haplogroup J and the Jewish Aaronic priesthood, which has an identified clear lineage to Shem, is at J2. This Haplogroup division identifies also the Buba clan of the Lemba tribe of Zimbabwe as Aaronic priests, and they have been separated from the rest of Judah/Levi for up to 2,500 years. Thus the J2 division is at least as old as that separation.
There are also a significant number of divisions in Judaism that show that Judaism is a religion and not a single Haplogroup lineage.
For example, whilst the Aaronic priesthood is identified as J2, the Levitical structure of Ashkenazi Jews are 52% R1a1, which is an identified Japhethite lineage occurring in Russia and the Central and Eastern Steppes and among the Aryans in India. It is Slavic. Also, 25% of all East European Jews are E3b, which indicates an Hamitic origin.
One hypothesis concerning the E3b origin is that Egyptians interbred with the Israelites. Another is that the Mixed Multitude involved E3bs, as there were some two million Israelites and approximately six hundred thousand of the Mixed Multitude. That would constitute 25%. However, when we read the genetic accounts of the Milesians they claim to have been part of the mixed multitude also in the Exodus. They are R1b. Thus, the explanation must involve later conversions to Judaism from Hamitic lineages. We find these in the occupation of Canaan under Joshua.
Canaan was a son of Ham and a number of sub-tribes were spared and joined Israel in some cases by subterfuge. The Ethiopian conversions were also of significance and so we have ample explanation for the E3b lineages at 25% of the Ashkenazim. The Amorites and the Southern Hittites also entered and bred with Israel. There are also significant levels of E3b in Syria, Turkey and among the Greeks at up to 30%. The Greeks are up to 30% Haplogroup J or known Semites also with up to 20% Hg I as well, which is also a prevalent Haplogroup among both known Semites and Europeans. We will deal with these aspects later.
Ashkenazi mtDNA
Another important aspect is that of the mtDNA of the Ashkenazi Jews. The mtDNA of the Ashkenazi Jews is classed as Near Eastern, but we need to look at the structure. As we saw with the mtDNA, all other groups came from Hg L3, which split into two main groups M and N. From N came R and the subsequent divisions of N and R. N split in the Middle East and formed the Nu subgroup of N (N1), I and W. We know that I occurred among the progenitors of the Kurds in what is now Kurdistan and the hill country on the borders areas of Iraq and Iran or Persia where the Medes (who are the sons of Madai) once lived. The other groups in this sub-group formed N1 etc.
10% of Ashkenazi Jewish mtDNA is N1b. This type differs only by a single transition from the ancestral N1b (145-176G-223). This may be among the most ancient of the mtDNA with the exception of L2. That L2 DNA as we have seen is an offshoot of the primal L mtDNA. It is of Hamitic origin and mostly now sub-Saharan and Ethiopian. It may have come in from the Canaanites or the mixed multitude. Moses’ first wife was an Ethiopian also and the Canaanites may well have had this DNA. There is also a Near-Eastern type of M1, which is the primal subdivision of the M group. The remainder are all subdivisions of R, and which constitute 60% of Ashkenazi mtDNA.
Near-Eastern mtDNA types among Ashkenazi Jews are N1b, H, J, K, L2, preHV, U7, M1, U1B (cf. Behar et al., Differential Bottleneck Effects in the mtDNA Gene Pool of Ashkenazi Jewish Populations, pp. 8,9,19; see also MtDNA Evidence for a genetic bottleneck in the early history of the Ashkenazi Jewish population, European Journal of Human Genetics, 2004, pp. 1-10).
There are low percentages of mtDNA such as U5a and V, which are considered to have come from European admixture (Behar et al., p. 10). However, V is predominant among the Saami with minor percentages at significantly less than 10% among the Europeans.
The latest division is that of U, which split to form K.
32% of all Ashkenazi-Jewish mtDNA is K, consisting of 4 major subtypes. Of the non-Jews the highest percentage is among the Druze at 16%. The next highest Ashkenazi-Jewish mtDNA is H at 21% and of which 7% is H ancestral. The British Ashkenazi-Jewish mtDNA is the CRS modal haplotype. Thus we might assume that the original division of HV is likely to have occurred on the early population of these females (early Israel?). J1 is next at 7%. (Behar op. cit.)
What is important in this aspect is that H constitutes 21% of Ashkenazi-Jewish mtDNA and that DNA is the major mtDNA sequence of the entire European population at just under 50% of Iceland, to up to 60% of the British Isles, and 65% of Iberia and 50% of France. It slowly decreases as the populations goes east. It drops to 25% among the Turks and Palestine/Egyptians and drops further among the Kurds, Persians, South Pakistanis and Hazara, with an increase to over 22% among the Uzbeks and up to 30% in northern Russia among the Komi.
It is thus reasonable to assume that the same females that were the basis of the Western Europeans were the basis for many of the Ashkenazi females, and that the mutations occurred within those tribes.
The basic mutation of U to K appears to have occurred in the Middle East in the area of northern Israel and the Lebanon, and included a significant portion of the females that formed the ancestry of the Druze. They were the ancestors of 32% of the Ashkenazi Jews. The other people with a high portion of K are the Kurds of which U forms the major portion. U forms significant portions of all European mtDNA. It is highest there among the Saami. HV, H, U, V and K are the main groups of all Europeans with I and J. H forms over 50% of all Anglo-Celts, Scandinavians and French and is even greater in Iberia. The significant difference between the Ashkenazi and the Europeans is that the division K is greater among them than it is in Europe and so the movement of K may have occurred later into Europe or been less in its mutation from U there. It is likely that these divisions occurred from the break-up of the Parthian Alliance and the movement of the Horde into Europe under Odin, commencing from the end of the second century CE. The two groups probably formed the same Scythian Horde comprised of Celts and Israelites and Jews from the Hittite Alliance. The Parthian and Scythian Horde moved into Europe and the other tribes formed the Khazar Horde and became the Khazar Empire, converting to Judaism ca. 740 CE, until the eleventh century and their defeat by the Russ and the later movement into Europe with the Huns. That explains why the non-Semitic Levite DNA in Ashkenazi Levites (which is the majority of its YDNA) is R1a1, which is Slavic rather than Celt.
We should also remember that Jewish identity since Talmudic times does not strictly follow the Bible system and has been acquired either by maternal descent from a Jewish woman or by rabbinically authorised conversion (see also Behar et al., Multiple Origins of Ashkenazi Levites: Y Chromosome Evidence for Both Near Eastern and European Ancestries, Am. J. Hum. Genet. 73:000-000, 2003). It is considered that the DNA came in from the Khazar conversion, and perhaps as many as 50 males originally enrolled as Levites from the Slavic Khazar Horde. There seems to be also a single male event that also expanded significantly after the movement into Europe (ibid.). Cohenim and Levites each comprise ~4% of the Jewish population. According to the Encyclopaedia Judaica (1972), Gentiles can become Israelites but not Levites or Cohenim (cf. ibid.). This rule obviously was not followed in the Khazar conversion. Within Ashkenazi Levite R1a1 haplotypes the microsatellite haplotypes are tightly clustered around a modal haplotype (16-12-25-10-11-13) that comprises 74% of Ashkenazi-Levites within the Haplogroup and 38% of Ashkenazi Levites overall. The haplotype is evenly distributed overall (Behar et al., ibid., see also Table 4).
Jews attempt to confine origins of the Ashkenazim to the small area of Northwest Europe from Northeast France to Northern Germany. They claimed an origin from as early as the 6th century. This area at that time was the central location of the Parthian Horde as it broke up and moved into Britain, Scandinavia, France and Spain and Italy. It is likely that some Jewish elements of the Horde moved there or followed the Horde later. The problem is that the earlier definitions, which saw Yiddish develop from the 10th century, was that the Ashkenazi language was ostensibly a language based on Hebrew letters but with words borrowed from the German lexicon. However, it is in effect a relexified Slavic tongue (Wexler, 1993, cf. ibid.). It is related to Sorbian and Belarus. This hypothesis is borne out now by the majority Slavic YDNA of Ashkenazi Levites. It is now considered to be Sorbian that was given Hebrew letters and a Germanic lexicography. The R1a1 YDNA also relate to the Sorbian and Belarus YDNA. These tribes might then be identified as the sons of Ashkenaz and part of the Khazar Horde, which is thus demonstrably Slavic.
The term Sephardi, when originally applied to Jews, was related to the Jews of Spain prior to the expulsion of 1492, and which Jews also were in Britain. However current usage applies it to all Jews of North Africa and the Near East who follow the Sephardi rite of worship.
The YDNA of the Levite Cohenim or priesthood, as opposed to the Levites and Israelites on the other hand, is a specific Haplogroup. The NRY (YDNA) of Ashkenazi and Sephardi Cohenim are demonstrably more related to each other than to other Israelites and non-Israelites (Skorecki et al., 1997; Thomas et al., 1998, cf. ibid.). The pattern of their DNA arises primarily from differences in the frequency of the Cohen Modal Haplotype (CMH), defined by six rapidly mutating microsatellites (Thomas et al., 1998), and a cluster of closely related haplotypes within a single Haplogroup (J2) defined by slowly mutating unique event polymorphisms. Chromosomes belonging to this haplotype and its related cluster were found at high frequency among Cohenim, but at a much lower frequency among “Israelites”. The pattern of cluster was found to be consistent with descent from a common ancestor who lived between 2,100 and 3,900 years ago. Now these are the findings based on the mathematical models that do not recognise faster YDNA mutation due to mtDNA exposure. Indeed, they would be limited in this haplotype also because of the limitations on the marriage of Cohenim outside of Levi.
The maximum earliest possible date for the
formation of the CMH would be 1900 BCE and thus Abraham could not possibly have
possessed this haplotype. Moreover, the lesser date, which is the most
probable, indicates that the CMH was fixed as late as the end of the Temple
period in 100 BCE. It may well be the result of the interaction in Judea
between the Edomites who were converted to Judaism under John Hyrcanus ca. 160
BCE and the Jews themselves there. That fact may well explain why the CMH is not
found extensively outside of rabbinical populations when we know for an
absolute fact that there was an extensive Levite population among the divisions
of the priesthood in Israel. All but three of these twenty-four divisions left
with the Israelite Horde when it was taken in to captivity by the Assyrians in
the eight century BCE and taken beyond the Araxes River into what became
Parthia and Scythia. There should be significant numbers of the original
Levite/Cohenim YDNA with the Horde wherever it is. The occurrence of the CMH among the Buba clan
of the Lemba in Zimbabwe indicates either that the mutation into the CMH and J2
was about the time of the Babylonian Captivity when the Lemba claim they became
separated and after the division of Israel into the ten tribes and the two
tribes of Judah and part of Levi and some of Benjamin, or their separation was
later and closer to 160 BCE when some of the priesthood moved to Egypt and
built the Temple there after the apostasy of Antiochus Epiphanes or with the dispersal
of Judea in the period ca. 135 CE after Hadrian’s edict and/or during the Bar
Kochba Revolt. The development from the Semite F of the Patriarchs to its
variants (e.g. G, I and J) may well have taken five hundred years or more in
the Middle East and in Scythia and occurred during the first millennium BCE or in the first century CE. The latest date is the most
probable for the dispersion, that is, under Hadrian.
We can thus deduce that the J Haplogroup was not the original Semitic haplotype. The Semites are thus of more than one standard Haplogroup in descent from Abraham. The original Semitic Haplogroup was F and divided into I and J at the very least, and probably G, I and J from the Middle East, in Parthia/Scythia and into Europe. An examination of this hypothesis can be conduced by referring to the results of Behar et al., Multiple Origins of Ashkenazi Levites ibid., Table A. In this table the Ashkenazi Levites were tested from 60 male lines over nine countries. These countries were Austria-Hungary (AH) x 10, Belarus (Bel) x 4, France (Fra) x 6, Germany (Ger) x 10, Lithuania (Lith) x 8, Netherlands (Neth) x 5, Poland (Pol) x 7, Romania (Rom) x 4, and Russia (Rus) x 6.
Of the 60 YDNA lines tested only eight were actual Semites. The other results were as follows:
Known Hamitic lines 12
DE (xE3) x 1 (Fra)
E3b x 11 (4 AH, 1 x Bel, 1 x Lith, 2 x Rom, 2 x Rus, 1 x Ger).
Assumed Japhethite lines 40
K (x L, N, O, P) x 1 (1 x Ger).
N (xN3) x 1 (1 x Lith).
Q x 1 (1 x Rus).
R1 x 1 (1 x Ger). This is a very rare line found in two Armenians previously.
R1a1 x 31 (4 x AH, 3 x Bel. 3 x Lith, 6 x Pol, 1 x Rom, 1 x Rus, 4 x Fra, 7 x Ger, 22 x Neth).
R1b x 5 (2 x AH, 1 x Rus, 1
x Fra, 1 x Neth).
The Semitic lineages are as follows and help demonstrate the point of the divergence of the Semitic lineages argued above.
F (x G, H, I, J,
K) x 1 (1 x Rom)
I x 1 (1 x Neth).
J (x J2) x 2 (1 x Lith, 1 x Rus).
J2 x 4 (2 x Lith, 1 x Pol, 1 x Neth).
The implications of these YDNA samples are quite far reaching. There are only 4 lineages that share the same J2 lineages as the CMH. Only 6 share the J Haplogroup. Thus only 10% of tested Ashkenazi Levites are recognised as of known direct Jewish/Israelite YDNA. F and I comprise 25% of the Semitic Levite YDNA tested.
Under current understanding, the staggering figures are that two-thirds of Ashkenazi-Levite YDNA is not Semitic. Almost 10 % are Celt and an amazing 52% are Slavic YDNA.
The studies conclude also from the homogeneity of the R1a1 modal haplotype that the ancestor was perhaps only 663 years ago. The growth may have been only from one person. This would have coincided with the establishment of the Jewish communities of Mainz and Worms (cf. Encyc. Judaica, 1972). The star-like patterns of the R1a1 YDNA indicate rapid population growth in the Ashkenazi Jewish population in the last 1000 years. The diversity in the Ashkenazi-Levites suggests a genetic intrusion after the Ashkenazi split from other Jewish populations (see Behar et al., ibid., and Table 5). The origin of the R1a1 Ashkenazi Levites is thus probably from Sorbian-speaking Khazars who converted to Judaism and entered into Levi and who then expanded rapidly in Europe.
Of the tests done on the Jewish populations, we can compare the Sephardic Cohenim and the Ashkenazi Cohenim and we find that even in the Cohenim there are Hamitic and Japhethite lineages (Behar et al., ibid., Table 1). Whilst the Haplogroup J has a frequency of .8684 for Ashkenazi Cohenim (AC) and .7536 for Sephardi Cohenim (SC), there is still a frequency for non-Semitic priests of between 14.16 and 24.64 per cent.
These are the non-Aaronic and indeed non-Israelite Cohenim:
Hamitic
BR (ex DE JR) at .0263 for AC and .0290 for SC.
There are E (ex E3a) at .0395 for AC and .0435 for SC.
Assumed Japhethite Cohenim
K (xL, N3, O2b, P) at .0263 for AC and .0290 for SC.
P (xR1a) .0263 for AC and .0870 for SC.
R1a1 .0132 for AC and .0580 for SC.
Of the basic Jewish populations only some 37% are actual Semites in the frequency samples (Behar et al., Table 1).
Hg J is for Ashkenazi Israelites (AI) .3700 and for Sephardi Israelites (SI) it is .3651.
Hamitic groups are:
Y (xBR,
A3b2) .0159 for SI
BR (x DE JR) .1400 for AI and .0794 for SI
E (xE3a) .2200 for AI and .1905 for SI
Assumed Japhethite lineages:
P (xR1a) .1500 for AI and .2063 for SI.
R1a1 .0400 for AI and .0159 for SI
Note there are far smaller percentages of Khazar converts among the basic Israelite stock than among the Ashkenazi Levites. The conclusions that might be drawn from these statistics are that the families of the nobility, and perhaps of the Kagan and the Beck of the Khazars, were converted and given Levite status, and/or that the converts who remained faithful to Judaism went west into Europe with the Huns, who were followed by the Bulgars. The paganised Slavs remained separate or behind to be swept away by the Russ and then the Mongols of 1215, and came in later as the Sorbians and the Belarus after the Huns and Poles.
What is certain is that the adherents of Rabbinical Judaism are not wholly Jewish or Israelite, and 63% of all Jews are either Hamitic or Japhethite converts to Judaism; that is, unless it can be shown that some aspects of K are Semitic rather than Japhethite, which would throw the YDNA sequencing into complete revision.
What is of interest in the K2 lineages is that of the 117 K2 Haplogroup males in the data banks of the University of Arizona, some 15% are Jews. K2 YDNA is of Middle-East origin. Dr. Wells is of the opinion that K2 YDNA is ancient Phoenician. That would mean that the Phoenicians were biblically of Japheth, but we know historically that they are of Edomites who mixed with Hamites. Thus, they are Semitic and Hamitic. The only Japhethite sea-trading nations allied with them were the ships of Tarshish and of the Kittim, the Dodanim and those of Javan. If K2 is indeed a major Phoenician YDNA, then it demonstrates that the Ancient Sea Kings were an aggregation of the trading alliances of the three ethnic subdivisions of the sons of Noah. This then reflected itself in the conversions to Judaism among the trading partners.
The Sons of
Shem
The sons of Shem are Elam and Asshur, Arphaxad and Lud and Aram.
We know for certain that the Elamites became the Persians, the Sons of Asshur became the Assyrians, the Sons of Arphaxad became the Hebrews, and Aram became the Syrians and the source of the name of the Aramaic language. They also later went north into Armenia.
From the modern DNA sampling of the Assyrian project, the modern Assyrians are L, G, J and some R1b1, but it seems that the R1b1 tested are English on the paternal side and Assyrian on the maternal side. Thus, Northern Aram (in the area of Armenia and Georgia) and Asshur developed the G Haplogroup, which is where it is found in greatest percentages.
Josephus gives us a clear indication of the sons of Aram when he says that of the four sons of Aram, Uz founded Trachonitis and Damascus, which is the country between Palestine and Celesysria, Ul founded Armenia and Gather the Bactrians, and Mesa the Mesaneans now called Charax Spasini. From this record the Semitic G Haplogroup would appear to come from Aram and Asshur. Josephus records that Abraham first settled with an army in Damascus (ibib., Whiston, tr., p. 32).
Josephus says of Joktan of the Hebrews that he had thirteen sons: Elmodad, Saleph, Asermoth, Jera, Adoram, Aizel, Decla, Ebal, Abimael, Sabeus, Ophir, Eulat, and Jobab. He says that these people inhabited Asia from the Indian River Cophen and the lands surrounding it. This is modern Kabul and the valley of its river. The land further towards the Indus is Bactria. Thus, if that is the fact of the matter, they seemingly moved through the Arabian Peninsula and most went on to India. The result could be the composite groups of Joktan in what is Afghanistan, Gather in Bactria and then Lud in the Punjab. The Semitic Haplogroup H in India would then be derived from a mutation that occurred among the Hebrews of Joktan and the Semites of Lud. The golden wedge of Ophir refers to the lands of one of the sons of Joktan who settled in Afghanistan or India, and thus the area of Ophir may be from the Hindu Cush into India or even Sri Lanka (Ceylon).
Afghan
and other Pashtun
The Wikipedia
article on Pashtuns says:
"According to most
anthropologists, Pasto-speaking Pashtuns appear to be primarily of Iranian
origin (as well as being modified by various other invaders and migrants over
the centuries) and are very similar to the Persians, Kurds, Tajiks and
Baluchis. These anthropologists believe the Pashtuns have eastern Iranian
origins as the Pashto language is classified as an eastern Iranian tongue
distantly related to Ossetic among other Iranian languages (see Ethnologue for
further details).
The Pashto-speaking
Pashtuns refer to themselves as Pashtuns or Pukhtuns depending upon whether
they are speakers of the southern dialect or northern dialect respectively.
These Pashtuns compose the core of ethnic Pashtuns who are predominantly an
Iranian people and found in southern and eastern Afghanistan and western Pakistan.
Many Pashto-speaking Pashtuns have however intermingled with various invaders,
neighboring groups, and migrants (as have the other Iranian peoples) including
possibly the Ghilzai who may have mingled with Turkic tribes, the Durrani who
have interacted considerably with the Tajiks (another Iranian people), and
Pashtun tribes north of Peshawar who have mingled with Dardic groups. In terms
of phenotype, the Pashto-speaking Pashtuns overall are predominantly a
Caucasoid people with Mediterranean features, but blonde hair and blue and
green eyes are not uncommon, especially amongst remote mountain tribes."
It also says that:
"From the 1st
century BCE to the 5th century CE the regions where the Pashtuns lived saw
immense migrations of Eurasian peoples including the Aryans, Persians, Sakas,
Scythians, Kushans, Huns, and Greeks. Later invaders would include Muslim Arabs
and Central Asian Turkic tribes as well as the devastating assaults by the
Mongols."
There would thus be
Japhethitic R1a and R1b and Hamitic C YDNA present.
The article says that
from:
"random sampling
of Pashtun populations in western Pakistan) the anthropological evidence that
the Pashto-speaking Pashtuns are an Indo-European people related to other
Iranian groups as well as speakers of Dardic languages such as the Kalasha and
the Nuristanis appears very probable but is not by any means conclusive at this
time. The testing, though still in its initial phases, has not shown any
substantial connection between the general Pashtun population sampled to the
genetic markers found amongst most Greeks, Jews, or Arabs. What may be the case
is that the Pashtuns have been slightly modified over time by various invaders,
while maintaining their eastern Iranian base genetically overall. Ultimately,
more research and a wider sampling of DNA will be required before the findings
can be deemed conclusive and generally representative of Pashto-speaking
Pashtuns."
The conclusions here
come from the premise that the Semitic Haplogroups are all J as found among the
three populations mentioned.
The Pashtun are
classified as Iranian, which as we have seen is primarily Haplogroup I, but
they vary as a typical Central Asian composite YDNA grouping. An attempt was
made to classify the Pashtun in the 16th century as one of the Bani
Israel, of the Lost Ten Tribes and as Joseph. This was debunked based on their
Indo-Iranian affinities and language. However, it is an historical fact that
the area of Khandahar, Kabul and Bactria also, on the other side of the
Hindu Cush, were made provinces or satrapies of the Persian Empire and
their languages are thus affected by that fact. The DNA similarities with
Elam and their reported historical descent and affinity with Joktan indicate
that the original DNA of the Semites including the Joktan Hebrews was not
J but F, and the primary Semitic derivative is I and the secondary derivatives
are J, and then H and G. All the Eastern Semites of Elam and Joktan were Hg I,
rather than J. Thus these mutations occurred over the second millennium BCE
and the extended mathematical models of the evolutionists are wrong.
The Durrany and Galzay tribes are held to be direct descendants of the Ibrani or Hebrews and probably of the sons of Joktan. Some also hold the Rabbani, Shinwari, Levani, Deftali, and Jaji from Afghanistan, and the Efridi and Yusufzai from Pakistan to also be Hebrew Tribes.
Lud
Lud is a source of uncertainty and there are two explanations for the identification and locations of the sons of Lud. We will return to this puzzle shortly.
Other
Locations of the Hebrews
The origin and location of the Hebrews is identified with Abraham as coming out of Ur of Chaldea, and the sons of Arphaxad are identified with the Akkadian groups in the Mesopotamian plain between the children of Asshur or the Assyrians and the Sons of Elam, who were on the east of the Tigris basin to the Persian plateau.
The other group of the Medes who were associated with the Persians and occupied the high country to the north of Elam and Gutea are not Semites – they are the sons of Madai, a Japhethite tribe and are identified today as the Kurds. We will return to them later. The Guteans or Guti also may well be associated with the Goths as part of the later Scythian Horde.
Professor Cyrus Gordon also holds the early Minoans to be Semites and their language Linear A as being Semitic. His work on Linear A has been met with controversy.
The sons of Arphaxad are Salah and Eber (from where the name Hebrew is derived). His sons Peleg and Joktan saw the split in the Hebrews. The sons of Joktan were thirteen in number and occupied the entire eastern area of the Arabian peninsula (Gen. 10:21-32) but many went on into Afghanistan or Pakistan, as we see above. The sons of Abraham through Hagar and some of the sons of Keturah, such as Midian, later joined them. The line of Abraham through Peleg was to Reu, Serug, Nahor, Terah to Abraham, Nahor and Haran. Abraham was called out of Ur of Chaldea and sent into the land of Canaan.
It follows that the lineages of the Semitic countries will give us clues as to the distribution of Semitic Haplogroups and mutations. The temptation is to say simply that Haplogroup J is the Semitic Haplogroup and all others are not Semitic, but that cannot be true given the distribution and mixtures of the nation groups.
For example, in the genetic structure of the Persians or Elamites the predominant is Hg I. It exceeds J by some 10%, and has an element of F and G with it. Those groups constitute half the YDNA of the Persians. The Middle-East Arabs also have a significant amount of I and some G. Haplogroups G, I and J constitute half of the Georgian/Armenians and the Turks, while F, G, I and J are half the Italians, with I and J comprising half the YDNA of the Greeks. When we go into Europe the balance of I increases dramatically.
G (M201), H (M69), I (M170, M258, and P19) and J (12f2.1) appear to be known Semitic lineages, with K (M9) the root base for all the sons of Japheth. Hg J decreases as it moves west in Europe but Hg I remains more or less highly significant as far west as the Anglo-Saxons and then decreases among the Basque, Welsh, Irish and Scots – but is nevertheless still found in quantities of up to 15% among these known Celts.
The Anglo-Saxons came in from the Middle East as part of the Horde at the fall of the Parthian Empire from what is now the area of Iraq and the area north of it. That occurred at the end of the second century CE. They moved in to the area of NW Europe and the Horde split up in various waves. The Anglo-Saxons and Jutes and the associated tribes of Lombards, Danes, Norwegians, Saxons and Fris, split up but retained a similar diversity. The northern French are also from the Middle East. The Normans constitute one element and are predominantly R1b but they contain other DNA groups such as I. The others are Francs in two groups. These are the Riphathian Francs and the Salien Francs. The Salien Francs could inherit through the male line only but the Riphathian Francs could inherit through both. As we know, Riphath was a son of Japheth, and we know that the nobility of these people also claimed descent from Antenor I King of the Cimmerians, and also from the Trojans. They were understood to be the offspring of the group that remained in the Middle East with the sons of Hector. They named their major cities after Paris and Troyes. Many of these people settled across the channel in Britain and those tribes (e.g. Parisii) were also R1b Celts. Over 35% of the Northern French are Haplogroups I and J. Approximately 35% of the Anglo-Saxons are Middle-East Hg I with some small amount of J. Approximately 40% of English are Middle Eastern and seemingly Semites. The majority of all NW Celts and Anglo-Saxons are R1b and part of the same genetic divisions as the known Japhethite tribes. The obvious exception is to the east where the Slavs are R1a, and the largest R1a divisions among the Horde coming into the North-West was among the Norwegians at about 30%.
Alliances
of Israel with Japheth
The historical works of the British tell us that the Welsh came in from the Middle East after the fall of Troy. They met up with the Cornish in North Africa, who were also survivors from the fall of Troy. We now know that it was the Trojan kingdom of Wilusia. Their histories say that they found sons of Magog already in Britain and subjugated them. They were sons of Riphath, and in the Middle East were part of the Hittite empire in alliance with the Hatti and Kalti further east.
The Welsh are known as the Kymry people and once occupied all of Britain and part of France as and with the Gauls. They were confined to Amorica, which is now Brittany. They were all Celts (Gimirra from Gomer). They are the origin of the Cimmerians (i.e. Gomerians) that reinvaded Anatolia and established Galatia. They formed Cleopatra’s bodyguard of 400 men that Caesar presented to Herod. They are probably the origin of the R1b in Turkey at >15% and to a lesser extent in Syria. The Turks are also a part of the Uralic-Altaic language group and so some may be R1a and R1b from that source also. These two groups and Q are very close and the division between the groups is jumped by very little mutation. Thus the American Indians are distant relatives of the Celts and Slavs.
Ireland
The early inhabitants
of Ireland were the Nemedians and then the Formorians, Firbolgs (or leather
people) and the Tuatha de Danaan.
The early descendants
came from Spain and were listed as Gadelians, the descendants of Gaodhal, sixth
from Japheth (op. cit.). The Gadelians were from the Scythians who extended
from the Boristhenes to the Massagetae (Greater Gothic Horde) and from the
Saces (hence Scythians) to the Caspian Sea. They were nomads (op. cit., p. 61).
Genetically we can
identify them as sons of Gomer and of Magog. The YDNA variations show that the
Ashkenazi Jews are more R1a1 than R1b and hence the variations must have
occurred after the divisions of the tribal groups. 52% of all Ashkenazi Levites
are sons of Gomer. Thus the prophecies regarding Gomer have direct relevance to
Israel today.
An ancient RxR1 YDNA
variety is found in Africa, specifically in Cameroon. It may have originated with tribal movements
of the Japhethite tribes into Africa, either from the East when the Australian
Aboriginal RxR1 groups left for Australia following the Cushite Hg C nomads
before them, or even as late as the Vandals who invaded North Africa from Spain
and were later defeated.
We know from Irish history and the genealogies that the Milesians were sons of Magog, and they test at R1b. However, we know they came into Ireland rather late and replaced a tribe that was there previously. That tribe ruled Ireland for hundreds of years and was known as the Tuatha de Danaan, termed the tribe of Dan or the Danaans. They are held to have also sailed with the Philistines, early Greeks and seemingly the Phoenicians (Danaans and Danites – Were the Hebrews Greek? BAR, June 1976). It is most likely that this tribe accounts for the incidence of Haplogroup I in Ireland as the Anglo-Saxons did not occupy Ireland when they entered Britain, while the Normans are predominantly R1b with some I from the Anglo-Saxons, but the percentage I would not proportionally represent the percentage in Ireland. The proportion of I is almost as high as the proportion of I in Scotland, but the predominant YDNA there also is R1b. There may be a percentage infusion of I from Norway in both areas. If the historical record and its interpretation that the Tuatha de Danaan are indeed an offshoot of Dan of Israel, then we have the demonstrable link that shows the Northern tribes as Haplogroup I. Thus 40% of the Anglo-Saxons would be Semites and probably of Israel, as the records assert. The argument against this aspect is that, of Jews, less than 2% are Haplogroup I. When we consider, however, that only 37% of Jews are Semites, that brings the sample up to some 6% of known Jews and thus may well represent the residue of the other tribes that remained with or returned to Judah.
There is also a known
incidence of Phoenician K2 from the ships of Tarshish that are present in Tyre,
Malta and in Wales. Thomas Jefferson, the US President, claimed Welsh descent
and was of this Welsh subclade of the K2 Haplogroup.
The first colony in
Ireland was commanded by Partholan, who was of the ancient line of the Greeks
before that land of Greece was invaded and populated by the E3b sons of Ham
from Africa. The second colony was commanded by Nemedius, who fought well to
consolidate his colony. However, the Formorians invaded and defeated Nemedius
in Cork and took over Ireland (MacGeoghegan and Mitchell, History of Ireland,
1867, p. 54).
The Firbolgs are
identified by MacGeoghegan as the Belgians and there was a tribe of Belgae in
Britain. The Formorians were either defeated or deserted the land and the
Firbolgs took possession of it. They were replaced by the Tuatha de Danaan, the
descendants of Nemedius who returned to Ireland through Germany and via Norway
and Denmark (MacGeoghegan, ibid., p. 55).
MacGeoghegan claims
that they may have been descended from the Dumnoniae of Cornwall. He states
that they were descended from Jobath, grandson of Nemedius, who led a part of
the colony into northern Germany from Ireland originally (ibid., p. 55). Thus
the original settlers of Ireland were an alliance of ancient Greeks and Tuatha
de Danaan or Tribe of the Danites, who were sons of Nemedius. These were the
early seafaring Danites allied with the Ancient Sea Kings.
MacGeoghegan also
claims that Briotan Maul, grandson of Nemedius by Feargus, established himself
with his tribe in Britain, which according to the Psalter of Cashel was
called according to his name – and they reportedly settled there as Britons.
MacGeoghegan says that this is at least as credible as the account by Geoffrey
of Monmouth. This would account for the I Haplogroup (Isles subclade)
distribution among the R1b Picts in Scotland. They did not come from Norway and
are almost exclusive to the British Isles. The Picts were also early
settlers in Britain virtually a millennia before the Scots.
At the same time,
either by the defeat or the departure of the Formorians, another group of 5000
men came in and divided Ireland into a Pentarchy, with Slaingey governor of
Leinster chief of the Pentarchy of Slaingey, Rory (Rughruighe), Gan, Gannan,
and Sengan – all children of Dela of the race of the Nemedians. The people were
then known by three different names: Gallenians, Damnonians, and Belgians. They
ruled for eighty years under nine kings, the last of which was Eogha. He
married the daughter of a Spanish prince. She was buried at Tailton in Meath,
which was named after her.
The Belgians or Belgiae were located nearest the Dumnonians in England also and it is likely that the three names were indicative of an alliance of the three tribes that moved from Europe to England and on to Ireland. The Gallenians were seemingly Gauls, hence also Galatians. However, it is likely that through the Nemedians under the Tuatha de Danaan groups that we see the Haplogroup I intrude along with the R1b (AMH) of the Celts. This group came from Denmark much earlier than the later R1b Danes of the Parthian Horde (the later Danes were DYS 390/391 23/11, and there are two I groups in Denmark also which differ, one being the Anglo-Saxon type; see below). The movement might also have been with the Huns or even prior to them from the shipping groups in the second millennium BCE. There were extensive amber and trading systems in the Baltic at that time. As seen above, we know that the K2 Tarshish/Phoenician traders were in Wales and Cornwall and that line still exists there.
It was from this time
that the Tuatha de Danaan descended on Ireland under Nuagha Airiodlamh, who
defeated the Firbolgs under Eogha in County Mayo. The Firbolgs were driven out
and sought refuge in the islands of the North (ibid., p. 55). These people who settled
in the Isles were R1b. Also, as we noted, there is the subclade of I1b2a–Isles
and this Haplogroup is found almost exclusively in the British Isles and
heavily in Scotland. It is not Norwegian Viking. The same I Haplogroup is found
among the Basques. It appears that it definitely did not come from Scandinavia
but rather the Tuatha de Danaan came in from the Middle East to Spain and some
remained among the Basques. The major element went on to Ireland and from there
went to Scotland and the Isles. They are found among the Picts also, so they
must have gone there before the Scots moved from Ireland to Alba, as Scotland
was called before Niall of the Nine Hostages renamed it. They did not come in
from Europe with the Scots or the Milesians. The significance of the Tribe of
Dan as the Tuatha de Danaan is that it is not confined to Ireland as many of
the British-Israelites thought. This group is concentrated in Scotland also
with smaller numbers among the Welsh, with some in England and perhaps among
the Basques. From Revelation chapter 7 we see that Dan and Ephraim cease to
exist as separate entities and the composite Joseph remerges, but this time
without Manasseh who is separate.
The Islands are those
of the coasts of Scotland as it was not for some centuries that any
colonisation of Thule or Iceland was made.
The Milesians came via
Spain to Ireland and a sub-tribe entered England from Spain and became known as
the Brigantes. They settled in West England in what is now roughly the area of
Lancashire.
The Irish – up until
the Milesians entered ca. 500 BCE, as we see above – were comprised of three
groups called the Formorians, the Firbolgs and the Tuatha de Danaan or Tribe of
the Danites.
The Milesians we know
definitely were R1b Celts and we have their genealogies and their DNA to match
those genealogies. These Celts accord with the other Celt R1b YDNA sequences
but differ according to regions and tribes.
The major determinant
factor in the R1b Celt Haplogroups in West Europe is found in the R1b1 haplogroup
divisions. Four subclades of R1b are derived from the repeats of the DYS
markers 390 and 391.
The Atlantic Modal
Haplotype (AMH), which has a value of 24/11 for 390/391, is found on the
Atlantic coast from Spain to Scotland. These Celts are those older inhabitants
of Europe and descend from Japheth through the earlier divisions and are found
among the Basques.
The Anglo-Saxons came
into Britain from after the Roman occupation of Britain in 410 CE and before
the Norman Conquest. Anglo-Saxons have both R1b and I Haplogroups. The R1b
(R1b1) subclade of the Anglo-Saxons is distinguished by the DYS 390 and 391
markers that have values of 23/11. This view is open to some discussion and may
not be a clear identification. These values are found in NW Europe among the
other nations of the Horde that established itself there in Flanders (Belgium),
Netherlands, NW Germany and Denmark. The Fris and the Anglo-Saxons both share
this identifying marker among others. They and the Saxons of Germany are
virtually the same people.
The other subclades of
R1b are repeats of 24/10, which are found throughout Europe, and repeats of
23/10, which are found in SE Germany and into Austria. Thus they are all from
different lineages of the Celts or sons of Gomer that came originally from
Scythia and Parthia.
The Haplogroup I
variants that are found among the Anglo-Saxons of I1a are I1a AS (1-8). I1a AS
is the most common variant of the I1 Haplogroup and is named because it is
found where the Anglo-Saxons originally settled in their move in from the
Middle East. It drops off as we move into south-east Germany and beyond. It is
probable that the variants of this Haplogroup mark the ancestral determinations
of some of the Ten Tribes. These nations are invariably interlinked with the
Hittite Celts as described above. The AMH is an earlier group distinct from the
Anglo-Saxons who had distinctive markers on both R1b1 and I1a AS.
The determination of
Isles I1a AS to the Danish versus the German ancestral accounts on the
continent will yield more information. The I1a- (Norse) is the most populous
form found in Sweden and Denmark and is close second in Norway. It is found in
only small quantities in Continental Europe south of the Baltic and North Seas
and takes second place to I1a –AS in Denmark. Thus the I groups mix in Denmark,
but the Danes are predominantly Anglo-Saxon types (see the paper Population
Varieties within Y-Haplogroup I and their Extended Modal Haplotypes (and also
R1b Varieties) by Ken Nordtvedt on www.northwestanalysis.net)
I1a uN (ultra Norse)
reaches its peak in Norway. It differs from I1a Norse by a shift at DYS 385 b
from 14 to 15. It also splits into two varieties based on the DYS 461 repeat
value. I1a uN1 has modals 12, 28 at DYS 461 and 449, which is common to most of
I1a. I1a uN2 has 11, 29 repeats at DYS 461 and 449 and is more strongly
associated with Norway over Sweden and Denmark. These divisions may have
occurred after the Great Migration period in Scandinavia from the third and
fourth century CE.
I1a1 (old I1a4) is a
small subclade of I1a found in about 10% of I1a from Eastern Europe.
I1b1 Din (Dinaric)
takes its name from a mountain range in the Balkans near where it is most
prolific. It has also spread through much of Eastern Europe.
I1b1 –West (Western) is
a variety of (old) I1b, P37.2 + is found in prolific amounts across Germany’s
Baltic and North Sea coastal areas and then spreads into the British Isles. It
is notably identified by 15 repeats DYS 388 instead of the usual 13 repeats of
Dinaric I1b1. This variety was also recently found in Estonia.
I1b1-Isles is a
recently discovered variety of I1b1, which seems to reside mainly in the
British Isles. Identification markers are DYS 385, 463, and 464.
I1b1a (old) I1b2, a
subclade of I1b1 M26+ is a large percentage of the population of Sardinia and
spreads into Iberia. It is not found in any significance in the North of
Europe.
I1b2a – Cont
(Continental) (old I1c) is the main variety of I1b2a. It is most dense in NW
Germany, Netherlands and up into Denmark, with some in southern Sweden and
Norway.
A significant amount is
found also in the British Isles from the Germanic and Scandinavia
immigrants/invaders.
I1b2a-Root is an
unusual variety of I1b2a that spreads from Italy and Iberia up into Denmark and
splits into two varieties based on four of the markers.
I1b2* is another
variety that is found from Italy and Iberia up into Denmark and also splits
into two varieties based mainly on the Marker DYS 448.
I1b* (old I1(y) is a
small variety within the tree and is 10/10 DYS 459a,b as one of its modal
features and 19.19 or 19.21 at YCA11a,b and 12,29 at DYS389i,ii.
It is considered that
the varieties of these I Haplogroup subclades will determine the Hebrew people
and the R1a and R1b divisions will determine the branches of the Hittite
alliance formed within Scythia and Parthia and the Steppes progressively into
Europe.
It is now being argued
and accepted that Haplogroups I and J are subgroups of F that were once a
combined IJ group, being one people.
The
Colonisation of Europe from the Middle East and Scythia
Israel went into
captivity under the Assyrians ca. 720 BCE and was taken into the North into
what became Parthia and Scythia. There they forged a union with the Celtic
people who formed the northern Hittites.
There were members of
the Hittite and Scythian horde that moved after the fall of Troy into Europe
and Britain and Ireland and elsewhere in Europe.
Under Odin, and after
their defeat by the Persians in Parthia in the second century of the current
era, the Parthian and part of the Scythian horde moved into NW Europe and
established a number of nations including the Anglo-Saxons, Jutes, Fris and
Dutch, Saxons of Germany, Danes, Norwegians and some of the Swedes, Lombards,
Goths, Suevi, Dalmatians, Vandals and others. They established the nations of
Western Europe, from Spain and Portugal to Denmark and Noway, across to Britain
and Thule (Iceland) and Greenland.
The Slavs were to move
in last, and thus the division is of West Europe under the R1b sons of Japheth
and East Europe under the R1a sons of Japtheth, all of whom are mixed with the
Hebrew Semites and the sons of Asshur.
The Slavs of Rosh,
Meshech and Tubal are all R1a, R1a1, Slavs and Scandinavians. The Russian
kingdom was established by the Varangian Russ of Scandinavia who established
kingdoms based on Kiev and Novgorod from 850-950 CE.
From Scythia to
Ireland
The Nemedians are the
progenitors of the Tuatha de Danaan. The timing of the occupation of Ireland is
of interest.
MacGeoghegan, reporting the ancient histories, says that Fenius Farsa, king of the Scythians had two sons, Nenual being the elder and heir to the crown.
The second was Niul,
who had become learned in the languages multiplied from the confusion at the
tower of Babel. Niul made a voyage into Egypt where he married Scota daughter
of the Pharaoh (Cincris) of Egypt and established himself in the land of
Capacirunt on the borders of the Red Sea. Niul had, by Scota, a son Gaodhal. He
was allegedly bitten by a serpent and presented to Moses at the time of the
Exodus. The bite turned green and the mark earned him the name Gaodhal-Glas or
Gadelas (glas meaning green in the Scottish language, so named
after Scota, his mother).
They were thus part of the mixed multitude in the Exodus. The Milesian genealogies appear to make this claim also.
They settled on the area of Egypt allotted to them by Pharaoh for his daughter’s progeny. After three generations they were attacked by the Egyptians under Pharaoh-En-Tuir and were thus forced to move, under their leader Sur, son of Easur, son of Gaodhal, to Crete where Sur died and was succeeded by his son Eibher, otherwise known as Heber-Scot (MacGeoghegan, ibid., pp. 56-57).
Heber-Scot moved from
there to the River Tanais in Scythia via the Aegean and Euxine Seas. They lived
there for four generations among the Scythians under Agnamon, Tait, Adnoin and
Lamphion.
The Scythians were as
far spread as the Uigur Autonomous region of what is now China in the second
millennium BCE. We have found their well-preserved mummies and tartan kilts,
many similar to Scottish hunting tartans.
The Gadelians were
suspected and persecuted and sought refuge among the Amazons under Adnoin, then
departed under his son Lamphion for Gaethluighe. Some think this to be Gothia,
but according to O’Flaherty it is Getulia in Africa, where they remained for
eight generations under eight kings, being the descendants of Lamphion; the
last being Bille father of Gallamh or Mileag-Espaine (in Latin) called
Milesius, who was ancestor of the Milesians or the ancient Irish. We now know
that the Amazons were married women of the Scythian tribes who fought with
their men from horseback.
Milesius returned for a
short time to Scythia where he was received by Riffloir, the king descended
from Fenius-Farsa through the elder son Nunnual. He appointed him general and
gave him his daughter Seaug in marriage, by whom he had two sons, Donn and
Aireach surnamed Feabhrua. On the death of his wife, and on falling out with
his father-in-law, he left with the boys and his troop of Gadelians for Egypt
where he received command of the force engaged in the war with the Ethiopians.
He was given a wife of
Pharaoh Nectonibus, also named Scota, by whom he had two sons, Heber-Fionn and
Amhergin.
In that time he had
twelve young men educated in the arts and sciences for the return to Spain.
After seven years in
Egypt he returned to Spain. After a great drought of several years the
Gadelians left for Ireland.
Thus we see the links
and the connections between the Scythians and the Irish and Scots. The
languages of P and Q Brythonic are different, but they are historically related
to the biblical personalities of Gomer and Magog.
Given the time-frame we
are also able to identify another YDNA link that appears to be the Nemedian or
Tuatha de Danaan. The Milesian Irish have a basic YDNA structure of R1b, which
is common to the Celts. It also has a YDNA Haplogroup that is I and appears to
be of Semitic origin.
This same Haplogroup is
also present in the Anglo-Saxons and the Europeans
generally in levels of up to 40%, as we see in
the Croats and the Norwegians also. Both these nations, however, have a major Slavic Haplogroup of R1a rather than the Celtic
R1b, which is nevertheless present in both groups.
By looking at the
histories and the DNA structure we are able to piece together the movement of
the tribes and make connections that would otherwise be impossible.
How would this Semitic
Hg I get into the Europeans?
The answer can be found
in the pages of the Bible.
There were the sons of
Keturah who moved into the North before the Exodus and who also became the
Spartan Greeks, as we know
from Josephus. These people were the people who moved on into Ireland, and part of this group may have formed the groups of the
Nemedians or the Tuatha de Danaan. The Tuatha de
Danaan are thought to have been associated with the Greeks, and it is possible that
the Danites associated with the sons of Keturah and hence they account for the I Haplogroup and also some of the J
Haplogroup of the Greeks. The E3b is a later intrusion from the Phoenician
Canaanites and from Africa.
These people also came
in waves from the Middle East. The Croats are also very high in the I Haplogroup
and as high as the Scandinavians in percentages. Thus throughout the Balkans we
have a high I Hg population from different sources but having much in common
ancestrally.
The
Hittite Alliance
From early in King
David’s career he established alliances with the northern kingdoms of Tyre and
Sidon and also with the Hittites, who were both sons of Riphath and sons of
Magog. They were weakened by the loss of the Wilusian kingdom whose capital was
at Troy. Thus the sons of Ashkenaz were exposed to Israel at this time and
Israelites joined them and later converted them.
Both the sons of
Ashkenaz and the sons of Riphath, and the sons of Togarmah, are all sons of
Gomer. Yet Magog also has R1b. Hence the divisions of R1a and R1b occurred
after the tribes had split. Some became R1a and some R1b. The sons of Ashkenaz
became R1a, or else the R1a groups called themselves Ashkenazi even though they
were Slavic, which is very possible from their Khazar exposure.
Saul consolidated the
area of Israel but David expanded it to the Euphrates.
He was in Cappadocia
and north of there into Scythia amongst Meshech and Tubal at the beginning of
the eleventh century BCE. We know this from the Psalms (Ps. 120:5).
Now there were two
Meshechs in the Bible. One was a son of Japheth; the other was son of Aram, son
of Shem, living in Syria. It is possible that David was in Syria among the sons
of Mesech, son of Aram, which had identified with the sons of Kedar.
The composite Meshech
and Tubal identifies the sons of Japheth and not the son of Aram.
The area of Cappadocia
in Turkey, which was taken over by Mesech millennia ago, was originally called
Kus after Cush the son of Ham and father of Nimrod. The king of the Hittite
empire had his capital there and the Babylonians referred to him as King of
Kus.
The Hittite empire was
divided into the northern and southern divisions. They were comprised of the
sons of Ham and the sons of Japheth, both of Gomer and Magog and Madai or the
Medes, and the trading alliances with them based on and with Sidon, from
Tarshsish and also the sons of Heth. Homer, in the Iliad, says that the
Trojan host spoke many languages.
The inhabitants of Tyre
and Lebanon were to become K2 YDNA descendants of the sea alliances of the sons
of Japheth (sons of Javan Tarshish and Elisha, and Kittim and Dodanim or
Rhodanim). These were not of the sons of Heth, and we see them in the
descendants of Lebanon and of Malta today.
The Haplogroup H is largely confined to India in both north and south and to a lesser extent north to the Uzbeks and Kyrgyz. The larger incidence in the south indicates that it was there before the Aryan invasion. If this is a Semitic Haplogroup we have to identify the Semite nation that mutated into this Haplogroup in isolation in India.
The Semitic tribe that would qualify is the tribe of the sons of Lud. They are not to be confused with the Ludim who are sons of Mizraim or Egypt. Josephus identifies these people with the Lydians that occupied the area of the Ionian coast in Anatolia or modern Turkey. That makes sense as they were then west of Aram and south of Asshur and Arphaxad among the Semite tribes and north of the Hamites. More is known of this section, but they and their descendants are not Hg H.
There was a tribe called the Lullubi in the area north-east of what is now Iraq that made war on the Akkadians. They are thought to be a clan of what are now the Kurds called the Lullu, but that would not be the same people as the body of the Kurds are sons of Madai of Japheth or the Medes – but they may well have been absorbed over that length of time.
Others, however, identified the sons of Lud as moving east of Elam. That might put them in the area of the Indus. The ancient city in the Punjab is Ludhiana, seemingly named after Lud. The explanation would thus be that the majority split and went east and established the cities of India.
It would also explain why the ancient cities of Harappa and Mohenjo Daro in the Indus basin had contacts with the Assyrians ca. 1750 BCE and adopted their systems of worship. The C Haplogroups we would expect to find there with the sons of Cush are outweighed by the other H, L and J groups in the south with the larger Aryan elements in North India of R1a, with the more ancient RxR1 of the Dravidians in greater numbers than R1a in the south. India is thus a combination of all three lineages, with the sons of Cush moving north and east.
The Sons of
Abraham
Abraham had a number of sons and also the tribes by Lot, his nephew, were associated with him. Lot founded Moab and Ammon in what is now Jordan. Isaac had two sons Jacob and Esau, and the sons of Esau founded Edom and became the Edomites or Idumeans that partnered the trading system with the Phoenicians. They later became converted to Judaism by John Hyrcanus in the era of the Maccabees after 160 BCE. They were absorbed into Judea and became the southern Jews as opposed to the Galileans.
Jacob had the twelve tribes: Joseph and Benjamin by Rachel; Dan and Napthali by Bilhah the maidservant of Rachel; Reuben, Simeon, Levi, Judah and Issachar were born to Leah; and Gad and Asher were born to Zilpah, handmaid to Leah. We know that of the sons by Leah the sons of Levi developed into J2 lineage (over 1700 years).
However, for the Irish element of the Tuatha de Danaan to be of Dan the tribe they would have to be Hg I. (Another explanation might be that they been completely eradicated and the Hg I elements are from Norway and England. The objection to this view is that the percentage of R1a would have to be as great as it is in Scotland, but it is minute in comparison. Thus the Hg I there is not from Scandinavia or England and is not Milesian either and hence is Tuatha de Danaan.) Many of the other sons of Israel also would have to be Hg I as well, and so the mutations would have had to occur not only among the wives but also among the sons of the same mothers. The other explanation is that the mutations occurred in Egypt and the tribes diverged there. The same divergences had to be occurring among the Joktan Hebrews and the Elamites and the sons of Keturah.
From the proportions of DNA among the Elamites and the Arabs, it appears that the earlier DNA developed independently among the sons from F into I as the predominant Haplogroup, but also into J and G.
It is recorded [cf. below] that the sons of Keturah were Zimran, Jokshan, Medan, Midian, Ishbak, and Shuah. Jokshan’s sons were Sheba and Dedan. The sons of Dedan were Asshurim and Letushim and Leummim. The sons of Midian were Ephah and Epher, Hanoch, Abidah and Eldaah.
We know that the Midianites were subjected to war firstly in the days before Israel went into captivity in Egypt and later after the Exodus due to idolatry. They were to occupy the area of Arabia with the sons of Ishmael and the Joktan Hebrews before them.
We know from the authority of Josephus that the Spartans were sons of Keturah. Thus the large amount of YDNA Hg I in Greece may be from the Sons of Keturah, with the J coming in later from the conquests in Asia Minor and from the Arabs in later centuries. There are significant Semitic Hg J populations around the Mediterranean but I is greater in incidence all over Europe. Greece also has the largest percentage E3b population outside of Africa.
If Haplogroup I is truly Semitic as it appears then the Bulgars and Romanians have the largest percentage of Semites in Europe. However, NW Europe and Scandinavia are not far behind them. The Anglo-Saxons carried with them when they came from the Middle East a calendar system based on the Semitic and it was called Al manach or “the counting” – and our word Almanac is derived from that system. An original 59-day bimonthly Almanac based on the New Moons is preserved at Cambridge.
It appears that the Parthian Horde was a composite of the Celtic and Semite horde. They had become one people of smaller tribes that moved and recombined to settle in the areas of Europe. Their language was Teutonic and they absorbed the Celts who had preceded them, but they themselves were a large percentage of Celt from the Hittite alliances they had formed in the Middle East.
Let us now look at the divisions that appear to be from the K Haplogroup from which the Japhethite tribes seem to be descended.
The Sons of
Japheth
Haplogroups K, L, M, N, O, P, Q and R appear to be Japhethite tribes. God said that He would bless Japheth. He was told that God would enlarge him and he would dwell in the tents of Shem. Thus we would expect to find a mixture of Japheth and Shem in the same national groupings. Thus Hgs I and R1b and R1a groupings satisfy these criteria.
K is the root division for all of the subsequent Haplogroup divisions from L to R2.
The majority of K is in Papua New Guinea and Melanesia and in the Han Chinese. There are smaller percentages among the Uygurs, Uzbeks and Mongols and among the Polynesians. Some K is also found among the Italians. Thus that root division moved progressively east, with a small amount remaining in the Mediterranean in Greece and Italy. The other groups mutated from that Haplogroup.
These are the sons of Japheth: Gomer and Magog, Madai, Javan, Tubal, Mesech and Tiras.
The sons of Gomer were: Ashkenaz, Riphath and Togarmah. These are the Gimirra mentioned by the Assyrians who were the Kimmerians of Herodotus. They are the progenitors of the Celts. The Ashkenazim formed the Khazar Horde. The Riphathians formed the Western or Trojan Celts of Wilusia. They became the ancient Britons from 1000 BCE, displacing or subjugating the Magogite Celts in Britain. The Anglo-Saxons and Jutes in turn displaced them from the fifth century of the Current Era. They repelled the Hun invasion of Britain in the 10th century when Humber king of the Huns was killed there at the Battle of the Humber River named after him.
Magog is associated with Gomer in Ezekiel 38:2,6. Magog is thus a progenitor of the Celts also, and they are the Milesians who later invaded Ireland in the fourth/fifth century BCE and the Scythian Celts who came in to Scotland in the fifth century CE. Josephus says that Magog was the progenitor of the Scythians (A of J, Bk. 1, ch. vi). The distinction in the Celtic language of these groups is termed P and Q Brythonic; they are not the same language. However, they came from Anatolia and the areas to the north in what was Scythia. The Hittite alliance had many languages and the Greeks describe the formations of the Trojan host as being of many languages.
We also know from the historical record in the UK that the Basques were part of the original inhabitants of the Iberian area of what is now northern Spain and are allegedly descendants of Godel. Godel or the Godelians are descended from Japheth. The histories also say that they are descendants of Tubal and of Tarsis (Tarshish), the son and grandson of Japheth (Catholic Encyclopaedia, Vol. XIV, p. 176). Tarshish was the son of Javan and not Tubal. Thus the histories hold there were two allegedly Japhethite groups in Spain. The Bible identifies the Tarshish (2Chr. 9:21; 20:36,37; Ps. 48:7; Isa. 2:16; 23:1,10,14; 60:9; Jer. 10:9; Ezek. 27:12,25; 38:13; Jonah 1:3; 4:2). These were the Iberians, who were divided into the Basques in the northern sector and the Tartesians in the south (ibid.). The Celts then superimposed themselves on these two earlier or aboriginal populations. Then the Celts were in the north and west and the Iberians in the east and south, with the fusion of the “Celtiberians” in the centre in Aragon and Castile. However, they too have an element of I among them, albeit smaller than that found in Ireland at >7% approx. The Phoenicians also established colonies there on the coasts at Cadiz, which was their chief city, and also at Carteya, Calpe, Malaga, and Sexi. The Rhodians also established a colony at Rosas south of Cape Cresus ca. 910 BCE, and the Phocians at Emporium. The Carthaginians settled in the Balearic Isles in the seventh century BCE. In the sixth century they aided the Phoenicians against the Tartesians and then took possession of the principal city of Cadiz and began trading in Beatica. They began conquering Spain after the losses of Sicily in the First Punic War but lost it completely after the Second Punic War.
The Basque language is held to be of the Uralic-Altaic language group and hence related to Finnish and the Asian Altaic, Mongol, Manchu-Tungus and Japanese. The Basques however are R1b and some I.
The comments above regarding the Iberians show that Tubal founded the Iberians, and Josephus says (ibid.) that they were the sons of Thobel – hence Tubal and Thobel are held to be the same. Originally the sons of Mesech were in Cappadocia (ibid.). The later Iberians were Western Goths (Visigoths) who came in with the horde in the second century CE.
We have identified the Medes as the modern-day Kurds, and Bullinger holds this to be the case. He says that they were the Amada or Amadai of the Assyrian monuments (ca. 840 BCE), and thus now the Kurdish tribes that were east of Assyria.
Togarmah is described as being in the uttermost parts of the North and would appear to be the Haplogroup N and its variations. The sons of Togarmah, were this to be the case, would be the M214 subdivision from K following the N split into N (LLY22). The Chinese and the Malays are O (M175). The Uralic-Altaic group and the Han Chinese would thus be sons of Japheth with a large mixture of Cushite Mongols of Haplogroups C and D and their females.
The sons of Javan were: Elishah, Tarshish, Kittim and Dodanim. The Bible then says by these were the isles of the Gentiles divided in their lands; every one after his tongue, after their families in their nations (Gen. 20:2-5).
It is these groups with the E3b Phoenicians of Tyre, Sidon and Carthage that formed the trading empire of the Ancient Sea Kings. The priesthood of the Cretans were lighter skinned than the others, as we see now from the reliefs at the palace at Knossos, and they appear to have also worshipped the Golden Calf, from the earrings in the museum on Crete. There is a significant percentage of E3b around the shores of the Mediterranean.
The sons of Javan were identified as the Ionians of the cuneiform and the Tel el-Armana tablets. The Greeks were first known by this name, but the modern DNA divisions of the Greeks indicate we are looking at a different people to the original Ionian Greeks.
The sons of Elishah are understood to be the Alasia of the Egyptians – in other words, the inhabitants of the Cilician Coast. Bullinger thinks that Tarshish is probably Tarsus. However, Tarshish was in Iberia or what is now southern Spain. It was from this port that most transatlantic commerce was undertaken in ancient times. The Kittim were progenitors of the ancient sea-trading empires run from Cyprus. The Dodanim are understood to be the Rhodanim (1Chr. 1:7, cf. Septuagint).
Meshech and Tubal were associated with one another. The Tabali are associated as and with Tubal. Meshech are associated with the Muska of the Assyrian monuments. We understand these as the Russians, and their Haplogroups are predominantly R1a with a small amount of R1b and significant N, I, and J, and some E3b in northern Russia.
These tribes were originally in the north of Asia Minor above the Magogites, but with the defeat of the Hittites and the fall of Troy they entered Anatolia in Cappadocia. This is the conflict referred to by King David when he is with Meshech (and perhaps Tubal). The tribes were much further south in Georgia/Armenia when he was writing ca. 1000 BCE. The area around Georgia was also termed Iberia from Iber. This may be the source of the name, and accounts for the use of the name in Spain.
No doubt David was trying to contain Meshech (and perhaps Tubal) as he mentioned that when he is for peace they are for war (Ps. 120:7). He also mentions the tents of Kedar. Ezekiel 38:2 refers to the leader of Ro’sh, Mesech and Tubal. Ro’sh means also chief or pre-eminent and thus it is translated Chief leader or prince rather than as a name; but it may be the origin of the term Russ. The Viking Russ from 850-950 CE formed the Varangian Kingdom of the Russ around Kiev and Novgorod. There are R1a elements in Sweden and Norway and much more so than elsewhere in West Europe, but the Slavs are the true R1a groups, with the greatest percentage among the Poles. The Ro’sh were associated with Russia and the cities of Muskva (Moscow) and Tobolsk were associated with the tribes. They moved north after their containment in the beginning of the first millennium BCE. Nevertheless, they also have a significant I Haplogroup population. When Ezekiel is referring to Gog and the Ro’sh prince of Meshech and Tubal he says they are in the lands of Magog. Anciently that area was north of Anatolia and east of the Trojan area in what is now Turkey, Armenia and Georgia.
Tiras was the original inhabitant of Thrace (from Tirasia), but the tribe descended from him does not appear to have remained there as the Haplogroups appear to be Semitic.
There is one Haplogroup that has developed from Haplogroup P (M45, M74, P27, 92R7, P69?): that is Q (M242, MEH2, P36). That is next to the Haplogroup R supergroup. Q occurs among the Selkups and Altai in Russia and then goes right in to Siberia and across the Pacific Ocean into America. There is one other way that the tribe could have moved and that is by sea across the Atlantic.
We know for a fact now that transatlantic shipping was occurring on a regular basis ca. 1000 BCE between Egypt and America. Recent forensic tests on the mummies in the museum in Berlin showed positive for cocaine and tobacco consumption on every mummy in the museum. The British Museum suspected the results were contaminated and tested all of their mummies for comparison and they found no cocaine in theirs, but every one was positive for tobacco consumption. Now these products grow only in America. The mummies were all around 1000 BCE or so. The trade appears to have gone on during the second millennium BCE down to the first millennium CE, as Cyrus Gordon shows from the evidence. The trade was across the Pacific and the Atlantic.
Perhaps the Amerindians moved as described above, but they certainly were trading across the Atlantic with the Iberian Tarshish and the Ancient Sea Kings at the time of King David and up until the Romans. There is evidence of Roman trade there in the first century of the Current Era. The Bar Kochba rebels allegedly also went there in the second century of the Current Era and their coins and graves have been found in the Midwest of the USA (see also professor Cyrus Gordon, Before Columbus, Touchstone, 1972).
One of the odd things about the transatlantic crossings is that there are tribes that reportedly appear to have links to the Welsh or ancient British, but our current records do not show the R1b Haplogroup in any quantities. These aspects need to be investigated further. The DNA of the various Indian tribes and especially those in Virginia and the north-east needs to be quantified, as Ogham script has reportedly been found there – and so the DNA should reveal contact if it has occurred. The YDNA tests conducted on North American Indians to date have revealed Q, Q3 and C3 only.
As we have seen already;
Haplogroup R has RxR1 basic from Africa to India to Australia.
R1a comprises the majority of the Slavs and Indo/Aryans while Haplogroup R1b comprises the bulk of the West Europeans, both Celts and Anglo-Saxon/Norman. R1a extends into Germany and Norway in greater quantities than found elsewhere in Western Europe. In Iceland we see R1a, R1b and I begin to merge with Q but only here (from Eskimo infusion there and on to some families in Norway).
If Haplogroup I is the definitive Semitic Haplogroup then the Semites are spread across Europe, mixed with Japheth. The Basques, Welsh and Irish are the more distinct single Haplogroup peoples (R1b) but even they have up to four separate Haplogroups, with the Basques and Welsh/Cornish having only two Haplogroups.
The argument might be raised that the Haplogroups of Shem and Japheth are both F and that all their sons are simple variations from them without distinction, and thus G to J are of both groups, with K forming the next division and one that does not appear in the known Semitic J. Thus K must include all Japheth, but it might be then argued that G to I is open to both. Those arguments do not appear to stand scrutiny on a simple population basis.
The Evolutionists argue that these lineages all crawled out of bogs with ancestors alive up to 130,000 years ago. Thus J is the Semitic lineage and the others are all independent evolutionary activities, but they admit it is all from a single ancestor. They argue that the Flood is a myth and that anyone who takes the Bible account seriously is simply ignorant of scientific reality.
For the Bible to be the inspired word of God it must explain the basic origin of the nations in a rational and true manner. What we have seen here is that the DNA of known tribal groups matches the DNA of other groups that are identified with their biblical relatives.
The other argument is that it is all simply happening much faster than we thought and that these variations occurred independently in multiple locations, and the DNA sequence is not indicative of the origins of the people. In the short term that hypothesis does not agree with known testing and historical examples.
What has been proposed here is in accord with known history and the Bible record. It provides an explanation using genetics that is explicable within the Bible record and is an acceptable alterative to evolutionary models using extended time-frames.
What is also possible is that Haplogroup I is the major Semitic Haplogroup covering the Hebrews and extending to the Northern Israelites and also to the Elamites. Whilst we have found no Haplogroup I among the modern Assyrians, rather only G and J with L, some of Hg I might have occurred anciently. The prophecy that Israel and Assyria will come hand in hand out of the north in the Last Days at the restoration may be because they are both dispersed in Europe and some may well be Hg I; but modern testing shows no Hg I in those groups identified as modern Assyrians. Haplogroup J is thus a later Middle-Eastern development in the Arab countries after the Babylonian captivity and covering Jews and Ishmaelites and Edomites absorbed into Judah. Even on Evolutionary models, which are very skewed to earliest possible dates BCE, the J2 Cohen Modal Haplogroup (CMH) is admitted to be a possible 2,100 years old. Thus Hg I could well have preceded it by centuries and identify the Ten Tribes of Israel, who are then spread throughout Europe. The blessing of Japheth places his descendants within the tents of Shem and thus prophecy is fulfilled and the blessings of Shem accrue to Japheth, even though Japheth is larger in percentage terms in each nation.
What is certain is that the sheer extent of the Haplogroups I and R1b and also R1a in Europe make it virtually impossible for nearly every person in Europe not to be a descendant of Abraham, Isaac and Jacob either though their maternal or paternal lines. Thus the Hittite alliance with Israel has made them one people spread over a vast area with various language entities. Abraham has indeed become the father of many nations.
Read also the following papers:
Sons of Japheth: Part I (No. 046A)
Sons of Japheth Part 1A: The Sons of HN (No. 046A1)
Sons of Japheth Part II: Gomer (No. 046B)
Sons of Japheth: Part III Magog (No. 046C)
Sons of Japheth: Part V Javan (No. 046E)
Sons of Japheth: Part VI Tubal (No. 046F)
Sons of Japheth: Part VII Meshech (No. 046G)
Sons of Japheth: Part VIII Tiras (No. 046H)
Sons of Ham: Part I (No. 045A)
Sons of Ham: Part II Cush (No. 045B)
Sons of Ham: Part IV Phut (No. 045D)
Sons of Ham: Part V Canaan (No. 045E)
Sons of Shem: Part I (No. 212A)
Descendants of Abraham Part II: Lot, Moab, Ammon and Esau (No. 212B)
Descendants of Abraham Part III: Ishmael (No. 212C)
Descendants of Abraham Part IV: Sons of Keturah (No. 212D)
Descendants of Abraham Part V: Judah (No. 212E)
Descendants of Abraham Part VI: Israel (No. 212F)
Sons of Shem Part VII: Charts for P212A-212F (No. 212G)
Descendants of Abraham Part
VIII: Thirteen Famines of Rebellion (No. 212H)